Distribución espacial de Vanilla bahiana (Orchidaceae) en dos fitofisonomías de restinga. El patrón espacial varía?
←
→
Transcripción del contenido de la página
Si su navegador no muestra la página correctamente, lea el contenido de la página a continuación

Collectanea Botanica 40: e001
enero-diciembre 2021
ISSN-L: 0010-0730
https://doi.org/10.3989/collectbot.2021.v40.001
Distribución espacial de Vanilla bahiana (Orchidaceae) en
dos fitofisonomías de restinga. ¿El patrón espacial varía?
FELIPE FAJARDO VILLELA ANTOLIN BARBERENA1,2, ESTÍBALIZ LÓPEZ
HERMOSO3,4 & MARIA APARECIDA JOSÉ DE OLIVEIRA4
1
Universidade Federal Rural da Amazônia, campus Capitão Poço, Herbário HCP, Estrada do Pau Amarelo s.n.,
Vila Nova, BR-68650-000 Capitão Poço, Pará, Brasil
2
Museu Paraense Emílio Goeldi, Coordenação de Botânica, av. Perimetral 1901, Terra Firme, BR-66077-830 Belém,
Pará, Brasil
Universidad de Alcalá, Plaza de San Diego, s/n, ES-28801 Alcalá de Henares, Madrid, España
3
4
Universidade Federal da Bahia, Instituto de Biologia, Barão de Geremoabo 147, Ondina, BR-40170-290 Salvador,
Bahia, Brasil
ORCID iD. F. F. V. A. Barberena: http://orcid.org/0000-0002-7298-2977,
E. López Hermoso: http://orcid.org/0000-0002-8039-5959, M. A. J. de Oliveira: http://orcid.org/0000-0002-8863-8951
Autor para correspondencia: F. F. V. A. Barberena (felipe.fajardo@ufra.edu.br)
Editores: J. López-Pujol & N. Nualart
Recibido 31 agosto 2020; aceptado 5 diciembre 2020; publicado on line 30 abril 2021
Abstract
Spatial diStribution of Vanilla bahiana (orchidaceae) in two reStinga phytophySiognomieS. doeS the Spatial pattern
vary?— Vanilla bahiana is an orchid restricted to Brazil, extremely ornamental, threatened with extinction and with
economic potential. We evaluated the spatial distribution pattern of V. bahiana in shrub and restinga forest formations
in a conservation unit in Northeast Brazil. The Morisita’s index of dispersion, the Morisita’s standardized index of
dispersion and the TTLQV method were used, and 1920 m2 were sampled, through the demarcation of six transects
of 40 × 4 m per phytophysionomy. Each transect was subdivided into 10 plots of 4 × 4 m. In the shrub formation, we
observed 49 individuals of V. bahiana and a greater abundance at the edges of the transects (plots 1 and 2). In the rest-
inga forest, we recognized 47 individuals of V. bahiana and a distribution more homogeneous way along the transects.
We found an aggregate pattern for the species in the shrub formation and a random pattern in the restinga forest. The
different patterns could be explained by differences in luminosity, but they could also be associated with the dispersal
of seeds or the presence of fungi of different species. Future studies with spatial distribution in Orchidaceae should be
multidisciplinary in order to investigate the main causes of the patterns on local scales. Specifically, understanding the
relationships between Vanilla species and mycorrhizal fungi may have positive impacts on the cultivation and commer-
cialization of vanilla.
Key words: Brazil; conservation; ecology; microdistribution; orchids; vanilla.
Resumen
diStribución eSpacial de Vanilla bahiana (Orchidaceae) en doS fitofiSonomiaS de reStinga. ¿el patrón eSpacial va-
ría?— Vanilla bahiana es una orquídea hemiepífita restringida a Brasil, extremadamente ornamental, amenazada de
extinción y con potencial económico. Se evaluó el patrón de distribución espacial de V. bahiana en las fitofisionomías
arbustiva y de bosque en un fragmento de llanuras costeras (restinga), en una unidad de conservación en el Noreste
de Brasil. Para ello, se utilizó el índice de Morisita, el índice de Morisita estandarizado y el método TTLQV, y se
muestrearon 1920 m2, a través de la demarcación de seis transectos de 40 × 4 m por fitofisionomía. Cada transecto se
subdividió en 10 parcelas de 4 × 4 m. En la formación arbustiva, se observaron 49 individuos of V. bahiana y mayor
abundancia en los bordes de los transectos (parcelas 1 y 2). En el bosque de restinga se contaron 47 individuos of V.
bahiana, pero distribuidos de manera más homogéna en los transectos. Se encontró un patrón agregado para la especie
en la formación arbustiva y un patrón aleatorio en el bosque de restinga. Los diferentes patrones podrían explicarse por
las diferencias de luminosidad, pero también podrían estar asociados con la dispersión de semillas o la presencia de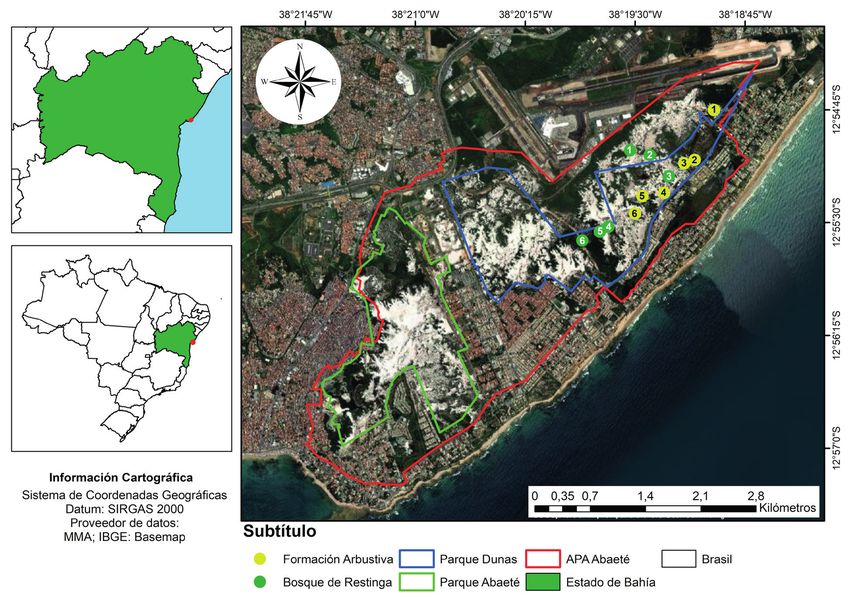
2 FELIPE FAJARDO VILLELA ANTOLIN BARBERENA, ESTÍBALIZ LÓPEZ HERMOSO & MARIA APARECIDA JOSÉ DE OLIVEIRA
hongos de diferentes especies. Futuros estudios con distribución espacial de Orchidaceae deberían ser pluridisciplina-
res con la finalidad de investigar las causas principales de los patrones en escalas locales. Específicamente, comprender
las relaciones entre especies de Vanilla y los hongos micorrícicos puede generar impactos positivos sobre el cultivo y
la comercialización de vainilla.
Palabras clave: Brasil; conservación; ecología; microdistribución; orquídeas; vainilla.
Cómo citar este artículo / Citation
Barberena, F. F. V. A., López Hermoso, E. & Oliveira, M. A. J. de. 2021. Distribución espacial de Vanilla bahiana
(Orchidaceae) en dos fitofisonomías de restinga. ¿El patrón espacial varía? Collectanea Botanica 40: e001. https://doi.
org/10.3989/collectbot.2021.v40.001
Copyright
© 2021 CSIC. This is an open-access article distributed under the terms of the Creative Commons Attribution 4.0 In-
ternational (CC BY 4.0) License.
INTRODUCCIÓN Souza-Leal & Pedroso-de-Moraes, 2014; Fajardo
et al., 2015; Rahal et al., 2015).
Los patrones de distribución espacial de los or- Los mecanismos de dispersión de semillas inter-
ganismos se pueden estudiar a diferentes nive- fieren primariamente en el patrón espacial y son co-
les, desde el nivel de región biogeográfica hasta rresponsables de los niveles de diversidad genética
el de la población (Hay et al., 2000; Tremblay et en Orchidaceae (Chung & Chung, 1999; Jacque-
al., 2006). La distribución espacial en microescala myn et al., 2007). Sin embargo, otros factores tam-
puede ser aleatoria, regular o agrupada y es deter- bién influyen en la distribución de las poblaciones
minada por factores abióticos (p. ej. disponibilidad de orquídeas, como las condiciones de microclima,
de agua, luz y nutrientes; temperatura y humedad principalmente humedad relativa, temperatura e in-
adecuadas) y bióticos (p. ej. competencia y her- tensidad de luz; el paisaje en sí; las características
bivoría), que interactúan entre sí, y son influen- morfológicas de la semilla; los sitios de dispersión
ciados por la acción antrópica y estocasticidad de propágulos; y la relación con hongos micorríci-
(Hutchings, 1997). La información sobre patrones cos (Budke et al., 2004; Phillips et al., 2010; Picco-
espaciales puede contribuir a una mejor compren- li et al., 2014; Rahal et al., 2015). Las especies de
sión de los procesos de dispersión de especies y re- orquídeas son particularmente vulnerables a la ex-
colonización de hábitats, así como para el estable- tinción, ya que, entre otros factores, con frecuencia
cimiento de estrategias de conservación y planes tienen poblaciones reducidas, con una distribución
de gestión, especialmente aquellos que involucren restringida, y dependen al menos parcialmente de
la recuperación de áreas degradadas o fragmentos los hongos, principalmente, con respecto a la ger-
de vegetación (Anjos et al., 2004; Bonet et al., minación y el desarrollo de plántulas (Batty et al.,
2005). Se han realizado evaluaciones del patrón 2001; Barros, 2007; Pandey et al., 2013). Además,
de distribución espacial para especies de diferen- están sujetas a colecciones depredadoras por colec-
tes familias de angiospermas, helechos y gimnos- cionistas y comerciantes, que alimentan el comer-
permas, presentes en dominios fitogeográficos o cio ilegal activo (Barros et al., 2007; Barberena et
biomas diversos en diferentes etapas sucesionales al., 2018).
(Hay et al., 2000; Anjos et al., 2004; Kanieski et En Brasil, un país megadiverso en especies de
al., 2012; Sousa & Cunha, 2018; Teixeira et al., plantas, las restingas han sufrido con la retirada
2019). En este sentido, las Orchidaceae han sido clandestina de arena, colección depredadora de es-
uno de los grupos botánicos más investigados; los pecies nativas, deforestación y la expansión inmo-
estudios se centran en especies terrestres y tipos biliaria (Britto et al., 1993; Souza et al., 2012). Va-
de vegetación del bosque atlántico, como el bos- nilla bahiana Hoehne es una orquídea restringida a
que caducifolio estacional, el bosque semideciduo Brasil, que presenta un elevado interés ornamental
estacional y llanuras costeras (restingas) (Budke et y se encuentra en los dominios fitogeográficos de
al., 2004; Krahl et al., 2010; Carvalho et al., 2012; la Caatinga, Cerrado y el Bosque Atlántico, donde
Souza et al., 2012; Dubbern et al., 2013; Piccoli habitan principalmente en las restingas (Ferreira et
et al., 2014; Quenzer & Pedroso-de-Moraes, 2014; al., 2017; Barberena et al., 2019; Flora do Brasil,
Collectanea Botanica vol. 40 (enero-diciembre 2021), e001, ISSN-L: 0010-0730, https://doi.org/10.3989/collectbot.2021.v40.001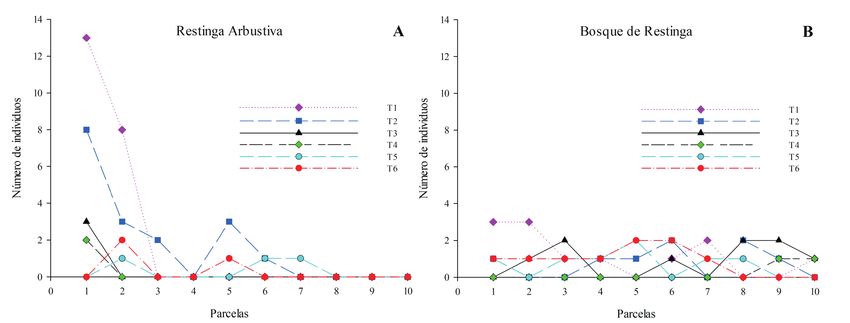
Distribución espacial de Vanilla bahiana 3
2020). La especie es indicadora de la etapa avanza- (Diário Oficial do Estado da Bahia, 1987). El clima
da de regeneración en fitofisiognomías arbustivas y de la región se clasifica como Af, definido como
de bosque de restinga en el estado de Bahía (MMA, cálido y húmedo, sin estación seca (Alvares et al.,
2012). 2013). La temperatura media anual y la precipita-
Recientemente, se ha sugerido el uso de V. bahia- ción media anual en la región superan los 25ºC y
na en los esfuerzos de mejora genética del género 2000 mm, respectivamente, y la humedad relativa
Vanilla Mill., ya que la especie es filogenética- varía entre 70-80% durante la mayor parte del año
mente cercana a Vanilla planifolia Jacks. ex An- (Viana & Kleinert, 2005). Las elevaciones del te-
drews, la vainilla más importante económicamen- rreno no superan los 100 m. El APA Abaeté cubre
te, y tiene varios atributos ecológicos deseables, dos parques municipales, el Parque do Abaeté, con
como gran cantidad de flores por racimo, extenso 270 ha, y el Parque Municipal das Dunas, con 290
período de floración, fructificación durante todo el ha, que es el fragmento de vegetación mejor con-
año y alta resistencia al estrés abiótico (Bouetard servado de la región (Diário Oficial do Município
et al., 2010; Anjos et al., 2017; Nascimento et al., de Salvador, 2011; Fig. 1).
2019). Además, V. bahiana produce cantidades Hasta ahora, las fitofisionomías del APA Abaeté
significativas de vainillina, sustancia extraída de no se han categorizado formalmente, pero las for-
los frutos maduros y que es ampliamente utilizada maciones Arbusto no Inundable (AR) y Bosque de
en la gastronomía mundial e incluso en medicina Restinga no Inundable (BR) son fácilmente reco-
(Bythrow, 2005; Lopes et al., 2019). Sin embar- nocibles y se han adoptado en el presente estudio.
go, V. bahiana ha sido evaluada como en peligro La formación Arbusto no Inundable se encuentra
de extinctión dado que la mayoría de las pobla- comúnmente en lugares ligeramente más altos, sin
ciones se encuentran en hábitats muy fragmenta- inundaciones periódicas o constantes, y consiste en
dos (Ferreira et al., 2017). El conocimiento sobre arbustos mezclados con bromelias terrestres y cac-
distribuciones en microescalas y particularidades tos que forman matorrales de diferentes extensio-
ecológicas de parientes silvestres de la vainilla nes y alturas, intercalados con extensas áreas abier-
es deficiente, incluso sobre V. bahiana, y necesa- tas de arena, el principal constituyente del sustrato
rio para que futuras acciones de conservación y en estas formaciones. La formación BR también es
programas de cultivo sostenible se establezcan de libre de inundaciones periódicas por la subida del
manera más adecuada (Nascimento et al., 2019; nivel freático o el desborde de lagunas perennes y
Watteyn et al., 2020). se puede caracterizar por un dosel de hasta 15 m
En ese contexto ecológico-conservacionista y y una capa evidente de materia orgánica sobre el
considerando también el potencial económico de suelo. Attalea funifera Mart. (Arecaceae) es una de
la especie, el presente estudio tiene como objetivo las especies más características de esta formación.
evaluar el patrón de distribución geográfica de V. Datos fitofisiográficos adicionales se pueden obte-
bahiana en dos fitofisionomías de restinga en una ner de Barberena et al. (2019).
unidad de conservación en el estado de Bahía. En
particular, buscamos responder ¿El patrón espa- Breve caracterización morfológica y ecológica
cial de una especie varía en fitofisionomías dife- de la especie
rentes del mismo tipo de vegetación, en la misma
zona? Vanilla bahiana se identifica fácilmente en la zona
por el hábito hemiepífito (germina en el suelo,
pero busca un soporte sobre el cual crece y even-
MATERIAL Y MÉTODOS tualmente pierde el contacto con el suelo), pero
también por un conjunto de características mor-
Área de estudio fológicas: entrenudos espaciados (4 cm o más);
hojas sésiles y oblongas; sépalos y pétalos verdes;
El estudio se realizó en el Área de Protección y labelo blanco, entero, con franjas amarillas lon-
Ambiental de Lagunas y Dunas de Abaeté (APA gitudinales y margen amarillo verdoso a amarillo
Abaeté), que comprende ca. 1800 hectáreas y está (Fig. 2). La especie es autocompatible, aunque de-
ubicado en el municipio de Salvador, Bahía, Brasil pende de las abejas del género Eulaema Lepeletier,
Collectanea Botanica vol. 40 (enero-diciembre 2021), e001, ISSN-L: 0010-0730, https://doi.org/10.3989/collectbot.2021.v40.001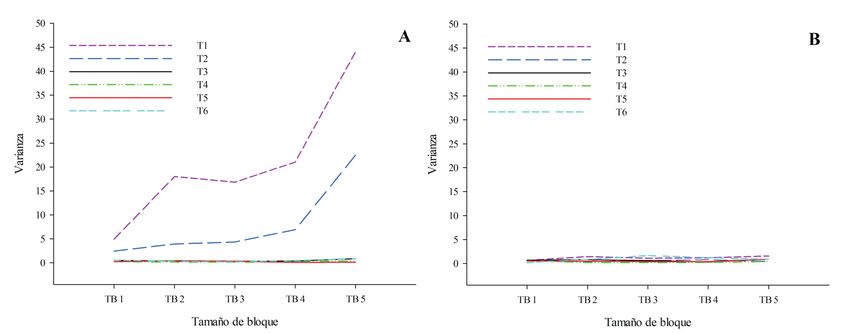
4 FELIPE FAJARDO VILLELA ANTOLIN BARBERENA, ESTÍBALIZ LÓPEZ HERMOSO & MARIA APARECIDA JOSÉ DE OLIVEIRA
Figura 1. Área de Protección Ambiental de las Lagunas y Dunas de Abaeté, Bahía, Brasil, destacando el Parque Municipal
Abaeté y el Parque Municipal Dunas. Los puntos amarillos y los puntos verdes corresponden, respectivamente, a los tran-
sectos establecidos en la formación arbustiva y en el bosque de restinga para evaluar la distribución espacial de V. bahiana
(mapa: José Antônio Lima Rocha Junior).
1841 (Apidae, Euglossinae) para su polinización y 331; Barberena & Aguiar 334) de acuerdo con las
reproducción, y presenta tasas bajas de fructifica- técnicas taxonómicas habituales y se depositaron
ción en la restinga (Anjos et al., 2017). Los indivi- en el herbario ALCB (Peixoto & Maia, 2013). Se
duos florecen y dan frutos durante la mayor parte muestreó un área total de 1920 m2, mediante el es-
del año y muestran un crecimiento exacerbado en tablecimiento de doce transectos de 40 × 4 m, seis
la unidad de conservación, llegando a decenas de en la formación de arbustos no inundable y otros
metros, aunque las partes vegetativas en contacto seis en la formación de bosque no inundable (Fig.
directo con el suelo y expuestas al sol están cla- 1). Los transectos fueron implantados en el Parque
ramente dañadas (Anjos et al., 2017; Barberena et Municipal das Dunas, la porción más conservada
al., 2019). de APA Abaeté, y sus alrededores (Fig. 1), con el
fin de minimizar posibles interferencias en el aná-
Recolección de datos lisis de los resultados, tales como la recolección
depredadora de orquídeas y los efectos asociados
Entre noviembre de 2014 y marzo de 2015, dos con la fragmentación de la vegetación. Se utiliza-
investigadores cuantificaron y recuantificaron ron fragmentos grandes para contener 10 parcelas
los individuos de V. bahiana de forma indepen- de 4 × 4 m separados a una distancia mínima de 50
diente, para evitar cualquier divergencia numéri- m entre sí, para evitar superposición. Sobre la base
ca (Barberena et al., 2019). Se recolectaron y se de estas premisas, el sistema de selección de las
herborizaron dos especímenes (Barberena et al. unidades de muestreo fue mixto, con la selección
Collectanea Botanica vol. 40 (enero-diciembre 2021), e001, ISSN-L: 0010-0730, https://doi.org/10.3989/collectbot.2021.v40.001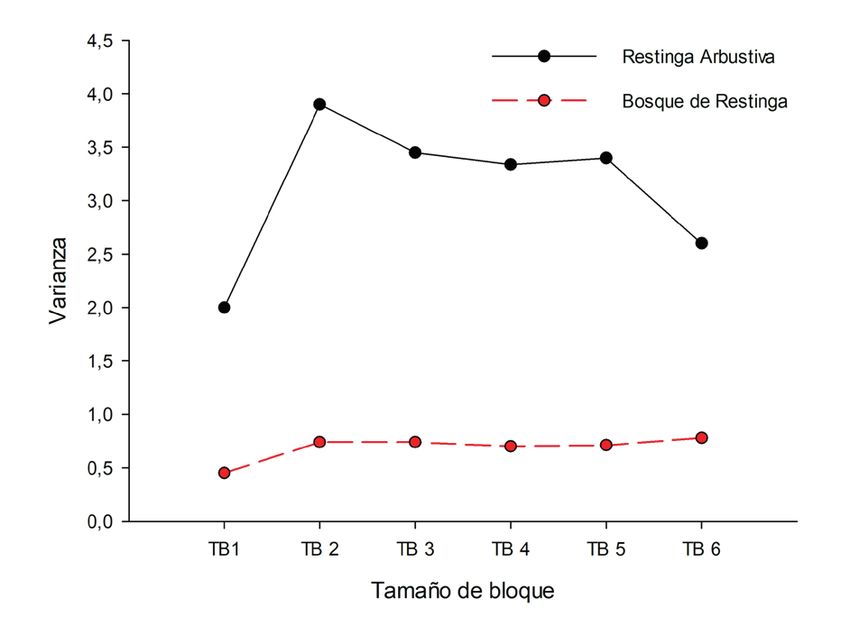
Distribución espacial de Vanilla bahiana 5 Figura 2. Vanilla bahiana: (A), hábito hemiepífito; (B), flor en vista frontal (fotografías: Felipe Fajardo Villela Antolin Barberena). aleatoria de unidades primarias (transectos) y el Para identificar el patrón de distribución es- establecimiento arbitrario de unidades secunda- pacial, se aplicó el índice de Morisita (Im), rias (parcelas). Las parcelas se distribuyeron con- que es considerado uno de los mejores índices tiguamente, siempre desde el área del borde hacia de dispersión y adecuados para el análisis con el interior del fragmento, contemplando un gra- transectos, y también el índice de Morisita es- diente ambiental (Krahl et al., 2010; Souza-Leal tandarizado (Ip), ya que es relativamente poco & Pedroso-de-Moraes, 2014). El criterio para la influenciado por el número de muestras y tamaño inclusión de individuos en la parcela fue el en- de las unidades de muestreo (Krebs, 2014; Oksa- raizamiento de la planta en el sitio (Souza et al., nen et al., 2019). 2012), por lo que los individuos que se extienden Para el Im, los valores estadísticamente signifi- a más de una parcela se contaron solo una vez. cativos >1 indican agregación, valores
6 FELIPE FAJARDO VILLELA ANTOLIN BARBERENA, ESTÍBALIZ LÓPEZ HERMOSO & MARIA APARECIDA JOSÉ DE OLIVEIRA
o iguales que 0,5 expresan agregación (con 95% RESULTADOS
de límites de confianza) (Oksanen et al., 2019).
De forma complementaria y para evitar estima- La distribución espacial de V. bahiana fue evalua-
ciones sesgadas, derivadas del uso de índice simi- da en dos fitofisionomías de restinga. Se contaron
lares y basados en la independencia de las muestras, 49 individuos de V. bahiana en la formación de ar-
también se utilizó el método de varianza móvil entre bustos (8,17 ± 8,49 individuos por transecto) y 47
bloques de parcelas (TTLQV), recomendado para individuos en el bosque de restinga (7,83 ± 3,31 in-
detectar la intensidad del patrón espacial (oscilación dividuos por transecto). En la fitofisiognomía de ar-
de densidad en la muestra) (Dale, 1999). La deter- busto hubo una mayor variación en la densidad de V.
minación del patrón de distribución se realizó a tra- bahiana entre los transectos, que fluctuó de 0,01875
vés de la interpretación del gráfico generado a partir individuos/m2 a 0,13125 individuos/m2 (3 a 21 indi-
del cálculo de las varianzas de conjuntos agrupados viduos por transecto), mientras que en la formación
de cuadrantes adyacentes. Si las varianzas fluctúan, de bosque la variación fue menor, de 0,0125 indi-
mostrando un pico en un tamaño de bloque (equi- viduos/m2 a 0,075 individuos/m2 (2 a 12 individuos
valente al radio del tamaño del grupo), el patrón es por transecto). De las 60 parcelas establecidas en la
agregado; si las varianzas estimadas son todas bajas AR, 14 presentaron individuos de V. bahiana (23,3%
y no fluctúan con el tamaño del bloque, el patrón es de frecuencia absoluta), con una clara concentración
uniforme; y si los valores de varianza fluctúan irre- de individuos en el borde de los transectos (parcelas
gularmente, el patrón es aleatorio (Krebs, 2014). 1 y 2), correspondientes a aproximadamente 82%
Aunque TTLQV asume la contigüidad de las del total de especímenes reportados para la fitofisio-
unidades de muestreo, se realizó un primer análi- nomía. En contraste, no se observaron individuos en
sis relativizando esta premisa, y considerando los las parcelas 8 a 10 en ninguno de los transectos de
transectos de cada fitofisionomía (y sus respectivas AR (Fig. 3A). En la formación de BR, los individuos
parcelas) como contiguos (en la secuencia de tran- se encontraron en 34 parcelas (frecuencia absoluta
sectos 1-6 para cada formación). Se adoptó el lími- del 56,6%), con una distribución más homogénea,
te superior recomendado del tamaño del bloque de habitando tanto parcelas en los bordes como en el
n
/10 (Krebs, 2014). Debido a que los transectos no interior de los transectos (Fig. 3B).
son realmente muestras independientes de la pobla- La varianza y la abundancia observadas resulta-
ción, se realizó también un análisis de TTLQV para ron en un patrón de distribución agregada para V.
cada transecto. Sin embargo, adoptamos n/2 como bahiana en la formación de AR y un patrón aleatorio
el límite superior del tamaño del bloque debido al en BR. Para la formación de AR, se encontró IM =
número reducido de unidades de muestreo. 1,82, con un estadístico de contraste χ2 = 44,18, muy
Figura 3. Abundancia de Vanilla bahiana por parcelas en el Parque Municipal Abaeté y sus alrededores, Bahía, Brasil: (A),
en la formación arbustiva; (B), en el bosque de restinga.
Collectanea Botanica vol. 40 (enero-diciembre 2021), e001, ISSN-L: 0010-0730, https://doi.org/10.3989/collectbot.2021.v40.001Distribución espacial de Vanilla bahiana 7
por encima del valor tabulado de 11,07. También se
obtuvieron Mu = 0,91, Mc = 1,16 e Ip = 0,57. El va-
lor de Im fue superior a 1 y el valor de Ip fue superior
a 0,5, resultados que indican distribución agregada.
En la formación de BR, se obtuvo IM = 1,04. Sin
embargo, el estadístico de contraste no difiere de lo
esperado al azar, y, por lo tanto, la distribución de
los individuos de V. bahiana en BR corresponde a
un patrón aleatorio. Esta inferencia fue confirmada a
través del Ip. Para la formación de BR, se encontra-
ron Mu = 0,91, Mc = 1,17 e Ip = 0,13.
Los resultados de TTLQV para transectos conti-
guos en la AR permitieron resaltar la gran oscilación
en la varianza (valores de 2 a 3,9) en relación con los Figura 4. Análisis de la estructura espacial de Vanilla
tamaños de bloque adoptados, con todos los valo- bahiana en el Parque Municipal Abaeté y alrededores,
res por encima del promedio de 0,82, y un pico muy Bahía, Brasil, según el método TTLQV y considerando
los transectos de cada fitofisionomía como contiguos: lí-
evidente en el tamaño de bloque 2 y otro pico menos nea negra contínua, en la formación arbustiva; línea roja
patente en el tamaño de bloque 5, que son indicati- discontínua, en el bosque de restinga.
vos de agregación (Fig. 4). Para la formación de BR
se obtuvieron valores de varianza menos discrepan-
tes, de 0,45 a 0,78, e iguales o cercanos al promedio con los tamaños de los bloques variaron poco, no
(0,78), sin la formación de picos, correspondiendo a excediendo de 0,9, y permanecieron cerca de los
una distribución aleatoria (Fig. 4). respectivos promedios de los transectos (Fig. 5A).
A su vez, el análisis de los resultados de TTLQV Así, es claro que el patrón agregado observado en
por transecto permitió detectar patrones distintos la formación arbustiva es el resultado de la mayor
dentro de la formación de arbustos. Hubo una agre- abundancia y distribución de individuos en los
gación clara en los transectos 1 y 2, representada transectos 1 y 2. Para la formación del bosque de
por una alta varianza (muy por encima del prome- restinga, sin embargo, se observaron patrones si-
dio para todos los tamaños de bloque) y picos muy milares para todos los transectos, marcados por la
claros, especialmente para los tamaños de bloque 4 fluctuación poco acentuada e irregular de los va-
y 5. Sin embargo, en los transectos 3 a 6 no se ob- lores de varianza, característica de la distribución
servaron picos y los valores de varianza en relación aleatoria (Fig. 5B).
Figura 5. Análisis de la estructura espacial de Vanilla bahiana en el Parque Municipal Abaeté y alrededores, Bahía, Brasil,
según el método TTLQV y considerando los transectos de cada fitofisionomía como unidades muestrales no contiguas: (A),
en la formación arbustiva; (B), en el bosque de restinga.
Collectanea Botanica vol. 40 (enero-diciembre 2021), e001, ISSN-L: 0010-0730, https://doi.org/10.3989/collectbot.2021.v40.0018 FELIPE FAJARDO VILLELA ANTOLIN BARBERENA, ESTÍBALIZ LÓPEZ HERMOSO & MARIA APARECIDA JOSÉ DE OLIVEIRA
DISCUSIÓN de la especie. Del mismo modo, principalmente
en la formación arbustiva, observamos parcelas
Se encontró que el patrón de distribución espacial con agrupación de individuos de V. bahiana (ocho
de una especie de orquídea puede variar en dife- o trece) y otras con pocos individuos (uno o dos)
rentes fitofisiognomías en la misma área. En la for- o incluso deshabitadas, ya que pueden constituir
mación de arbustos, verificamos el patrón agrega- manchas desfavorables, proporcionando un equili-
do para V. bahiana, pero en el bosque de restinga brio entre la colonización de hábitats disponibles y
encontramos el patrón aleatorio. Dichos patrones extinción de subpoblaciones locales (Tremblay et
fueron los mismos para los tres índices empleados. al., 2006).
Lima-Ribeiro & Prado (2006) demostraron para Uno de los requisitos para la germinación en Or-
especies de Annonaceae y Asteraceae que la adop- chidaceae está estrictamente relacionado con la
ción de diferentes métodos (sea los que utilizan el formación de asociaciones simbióticas con hongos
número de individuos por parcela o aquellos que micorrícicos (predominantemente especies de Ba-
estiman la varianza entre diferentes combinaciones sidiomycota) desde el comienzo de su ciclo de vida
de parcelas) para el análisis del grado de agrupa- (Feuerherdt et al., 2005; Dearnaley, 2007). Porras-
miento de una población no resulta en patrones Alfaro & Bayman (2007) demostraron que el géne-
espaciales distintos. Por lo tanto, los resultados ro Vanilla es generalista para hongos micorrícicos
encontrados en el presente estudio pueden consi- y que los hongos asociados con la germinación en
derarse consistentes. Además, varios autores han Vanilla son especies diferentes de las que ocurren
observado fluctuaciones (aumento o disminución) en otras etapas del ciclo de vida de las orquídeas de
en los valores de Im al cambiar el tamaño de las este género. Estos autores también observaron que
parcelas de ocurrencia de orquídeas terrestres, tan- la presencia de hongos micorrícicos en las raíces de
to para individuos inmaduros (que no muestran evi- especies de vainilla en el suelo es mayor que en las
dencia de actividad reproductiva durante el período raíces en la corteza de los árboles y que el género
de observación) como maduros (que muestran evi- Xenasma Donk (Ceratobasidium D. P. Rogers) es
dencia reproductiva), pero siempre sin alteración el más común en raíces en el suelo y el único en
del patrón observado (Budke et al., 2004; Piccoli et promocionar la germinacion de semillas de vaini-
al., 2014; Souza-Leal & Pedroso-de-Moraes, 2014; lla in vitro. En el suelo y en los forófitos, los hon-
Pedroso-de-Moraes & Souza-Leal, 2015). gos micorrícicos también tienen una distribución
El patrón agregado encontrado para V. bahiana en agregada y, aunque tienen una distribución inde-
la formación AR es repetitivo en Orchidaceae en el pendiente de la presencia de orquídeas (Feuerherdt
Neotrópico y se observa tanto en individuos inma- et al., 2005; McCormick & Jacquemyn, 2013), se
duros como en maduros de la misma especie y tam- encuentran poblaciones más grandes de hongos
bién en áreas de muestreo de diferentes tamaños (p. cerca de tales plantas (Perkins & McGee, 1995;
ej. Brudke et al., 2004; Otero et al., 2007a; Carval- Batty et al., 2001; Diez, 2007). Como el suelo es
ho et al., 2012; Souza-Leal & Pedroso-de-Moraes, heterogéneo en ambientes forestales se supone que
2014; Rahal et al., 2015). El patrón agregado se la disponibilidad de sustrato para hongos también
encontró para varias orquídeas terrestres en los ti- varía a microescala (Diez, 2007). Por lo tanto, la
pos de vegetación del bosque atlántico, incluidas disponibilidad de hongos compatibles es un factor
las formaciones de arbustos y bosques de restinga determinante para la presencia, abundancia y dis-
(Krahl et al., 2010; Souza et al., 2012). Adicional- tribución de orquídeas en un determinado lugar.
mente, Carvalho et al. (2012) también reconocie- Este aspecto podría explicar la variación entre los
ron el patrón agregado para V. bahiana en un frag- patrones de distribución de V. bahiana en las dos
mento de Caatinga en el estado de Bahía, aunque el formaciones de vegetación.
número de individuos observados fue bajo (n = 10). Las especies de Vanilla poseen semillas recalci-
La agrupación es favorecida por la presencia de trantes y crustosas con endospermo, y posible dis-
uno o más recursos favorables para el desarrollo de persión zoocórica; abejas, murciélagos frugívoros y
los individuos y la restinga es un tipo de vegetación aves son sugeridos como agentes dispersantes de se-
heterogéneo con parches apropiados (vegetación) millas en el género, además del viento asociado a la
y no apropiados (arena desnuda) para la ocurrencia gravedad (Cameron & Chase, 1998; Householder et
Collectanea Botanica vol. 40 (enero-diciembre 2021), e001, ISSN-L: 0010-0730, https://doi.org/10.3989/collectbot.2021.v40.001Distribución espacial de Vanilla bahiana 9 al., 2010; Soto-Arenas & Dressler, 2010). Aunque se influencia de las interacciones bióticas, como la produce la maduración del fruto de V. bahiana en la competencia inter e intraespecífica, herbívoros y APA Abaeté (Anjos et al., 2017; Nascimento et al., patógenos, y con el reclutamiento a una distancia 2019), no se observan plántulas y la forma de dis- más espaciada de la planta madre (Souza et al., persión de semillas en V. bahiana sigue siendo des- 2012). Por lo tanto, los diferentes patrones encon- conocida. Contrariamente a la percepción común, trados para V. bahiana podrían explicarse por dife- la dispersión de semillas de orquídeas por el viento rencias de luminosidad en las fitofisonomías, estar ocurre en distancias cortas de menos de 10 metros relacionados con agentes dispersantes de semillas o (Murren & Ellison, 1998; Chung et al., 2004). Ade- con la presencia de individuos de orquídeas en di- más, la dispersión de semillas por el viento a partir ferentes etapas del ciclo de vida, influenciados por de áreas abiertas hacia áreas de vegetación más ce- la presencia de hongos de diferentes especies. Así, rradas puede ser difícil (Azevedo et al., 2007), como los individuos de V. bahiana de las formaciones fo- a partir de la formación arbustiva, y puede contribuir restales pueden estar en una etapa más avanzada a generar el patrón aleatorio observado en los bos- de desarrollo que los individuos de las formacio- ques de restinga. Igualmente, la dispersión de semi- nes de arbustos, presentando un patrón aleatorio, llas por animales en la formación de BR puede ser resultante de la mayor influencia de las interaciones ineficiente para la formación de nuevos rodales de bióticas y reflejando una homogeneidad ambiental. individuos, ya que la circulación de dispersores en Independientemente de los factores responsables sitios más cerrados es más difícil. de estos patrones, nuestros hallazgos permitieron Además de la dispersión de semillas y de las rela- definir áreas de mayor y menor densidad poblacio- ciones micorrícicas, otro factor que probablemente nal y detectar discontinuidades en la distribución a interfiere con la distribución de V. bahiana en la pequeña escala de V. bahiana, lo que nos permiti- restinga es la luminosidad. Los hongos micorríci- rá adoptar estrategias de refuerzo de la población cos son más eficientes en la colonización cuando más detalladas para la especie en la APA Abaeté y son expuestos a alta intensidad de luz (igual o ma- posiblemente extrapolarlas a áreas de restinga ad- yor a 600 μmol m-2s-1; Moratelli et al., 2007), lo yacentes. que, a su vez, podría explicar las distintas micro- distribuciónes de V. bahiana en las dos fitofisiog- nomías. En la formación AR, se encontró un mayor CONSIDERACIONES FINALES número de individuos en los bordes de los transec- tos (parcelas 1 y 2), una región de luminosidad más Varios autores indican que el patrón de distribución intensa, con menor cobertura vegetal, mientras que, espacial en Orchidaceae se relaciona primaria- en BR, no se observó una mayor abundancia en las mente con la distribución y abundancia de hongos parcelas fronterizas o interiores (Fig. 3). El dosel micorrícicos, aunque la distribución de orquídeas heterogéneo de AR proporcionaría una mayor lu- también pueda ser determinada por factores adicio- minosidad en algunas parcelas, posiblemente cerca nales (Feuerherdt et al., 2005; Otero et al., 2007b; del óptimo para la eficiencia de la colonización por Waterman & Bidartondo, 2008; McCormick & Jac- hongos. Aunque no se obtuvieron los datos abióti- quemyn, 2013). De manera similar a lo verificado cos para cada parcela, la mayor presencia de indi- para las orquídeas terrestres, la especificidad de las viduos en las parcelas de borde asume que la luz (y orquídeas epífitas por ciertos forófitos puede ser un posiblemente la humedad) pueden explicar la dis- reflejo de la asociación entre hongos y especies ar- tribución en puntos de V. bahiana en la fitofisiono- bóreas (Otero et al., 2007a; Gowland et al., 2011) y, mía de arbustos. En contraste, el dosel homogéneo por lo tanto, la distribución del hongo sería el factor de BR resultaría en una menor penetración de luz limitante para la distribución de las orquídeas epífi- y diferencias de luz menos significativas entre las tas y no el forófito en sí. A su vez, la distribución y parcelas, haciendo que los microclimas en esta fito- abundancia de hongos micorrícicos también puede fisionomía sean similares. depender de factores bióticos y abióticos, incluyen- El patrón de distribución espacial en Orchidaceae do variaciones en microescala en las condiciones cambia según las etapas ontogenéticas, pasando de edáficas o de sustrato, lo que hace que la compren- agregado a aleatorio o uniforme, bajo una mayor sión de las causas de los patrones espaciales de las Collectanea Botanica vol. 40 (enero-diciembre 2021), e001, ISSN-L: 0010-0730, https://doi.org/10.3989/collectbot.2021.v40.001
10 FELIPE FAJARDO VILLELA ANTOLIN BARBERENA, ESTÍBALIZ LÓPEZ HERMOSO & MARIA APARECIDA JOSÉ DE OLIVEIRA
orquídeas sea aún más compleja (Osorio-Gil et al., Atlantic Forest. Nordic Journal of Botany 36: e01728.
2008; McCormick & Jacquemyn, 2013). Cierta- https://doi.org/10.1111/njb.01728
Barberena, F. F. V. A., Sousa, T. S. & Rocha Junior, J. A. L.
mente hay mucho que conocer sobre la actividad 2019. Mapping threats to the orchid populations in an en-
de forrajeo del hongo en habitats naturales y sobre vironmental protection area in Bahia, Northeast Brazil.
métodos de almacenamiento y cultivo de hongos Oecologia Australis 23: 346–356. https://doi.org/10.4257/
oeco.2019.2302.12
micorrícicos, así como sobre los transportadores Barros, F. 2007. A família Orchidaceae na Flora do Estado de São
de nutrientes y genes relacionados con el mante- Paulo e suas espécies ameaçadas. In: Mamede, M. C. H., Sou-
nimiento de la asociación simbiótica (Batty et al., za, V. C., Prado, J., Wanderley, M. G. L., Barros, F. & Rando, J.
G. (Eds.), Livro vermelho das espécies vegetais ameaçadas do
2001; Dearnaley, 2007).
estado de São Paulo. Instituto de Botânica, São Paulo: 47–52.
Bajo este sesgo, los estudios futuros con distri- Batty, A. L., Dixon, K. W. & Sivasithamparam, K. 2001. Con-
bución espacial en Orchidaceae en microescalas straints to symbiotic germination of terrestrial orchid seed
deberían ser pluridisciplinares, centrados en la in- in a Mediterranean bushland. New Phytologist 152: 511–
520. https://doi.org/10.1046/j.0028-646X.2001.00277.x
vestigación de las causas de los patrones y desaro- Bonet, A., Gras, M. J. & Raventós, J. 2005. Análisis de los
llados en línea con los estudios de la distribución de patrones espaciales de distribución de Vella lucentina M. B.
los hongos micorrícicos, ya que estos podrían tener Crespo para la selección de áreas de la Red de Microrreser-
vas de Flora de la Comunidad Valenciana. Mediterránea:
influencia directa en la comprensión de la biología, Serie de Estudios Biológicos 18: 8–43.
ecología, estrutura genética y acciones de conser- Bouetard, A., Lefeuvre, P., Gigant, R., Bory, S., Pignal, M.,
vación o restauración que involucren las orquídeas, Besse, P. & Grisoni, M. 2010. Evidence of transoceanic
siendo especialmente importante para especies ra- dispersion of the genus Vanilla based on plastid DNA. Mo-
lecular Phylogenetics and Evolution 55: 621–630. https://
ras, amenazadas de extinción o endémicas como V. doi.org/10.1016/j.ympev.2010.01.021
bahiana. Específicamente, comprender las relacio- Britto, I. C., Queiroz, L. P., Guedes, M. L. S., Oliveira, N. C. &
nes entre especies de Vanilla y los hongos mico- Silva, L. B. 1993. Flora Fanerogâmica das Dunas e Lagoas
do Abaeté, Salvador, Bahia. Sitientibus 11: 31–46.
rrícicos puede generar impactos positivos sobre el Budke, J. C., Giehll, L. H., Athaydel, E. A. & Záchial, R.
cultivo y la comercialización de vainilla. A. 2004. Distribuição espacial de Mesadenella cuspidata
(Lindl.) Garay (Orchidaceae) em uma floresta ribeirinha em
Santa Maria, RS, Brasil. Acta Botanica Brasilica 18: 31–
35. https://doi.org/10.1590/S0102-33062004000100004
AGRADECIMIENTOS Bythrow, J. D. 2005. Vanilla as a medicinal plant. Seminars in
Integrative Medicine 3: 129–131. https://doi.org/10.1016/j.
Agradecemos al Sr. J. Santana, gerente del Parque Municipal sigm.2006.03.001
das Dunas, por la autorización otorgada para realizar este estu- Cameron, K. M. & Chase, M. W. 1998. Seed morphology of
dio, y a J. A. Lima Rocha Junior, por la elaboración del mapa vanilloid orchids (Vanilloideae: Orchidaceae). Lindleyana
y de los gráficos. 13: 148–169.
Carvalho, D. N., Berg, C. van den & Pigozzo, C. M. 2012.
Floristic and spatial distribution of Orchidaceae species in
the Serra do Mucambo, Conceição do Coité, Bahia, Brazil.
REFERENCIAS Brazilian Journal of Ecology 14: 66–73.
Chung, M. Y. & Chung, M. G. 1999. Allozyme diversity and
Alvares, C. A., Stape, J. L., Sentelhas, P. C., Gonçalves, J. L. population structure in Korean populations of Cymbidium
M. & Sparovek, G. 2013. Köppen’s climate classification goeringii (Orchidaceae). Journal of Plant Research 112:
map for Brazil. Meteorologische Zeitschrift 22: 711–728. 139–144. https://doi.org/10.1007/PL00013868
https://doi.org/10.1127/0941-2948/2013/0507 Chung, M. Y., Nason, J. D. & Chung, M. G. 2004. Spatial genet-
Anjos, A., Mazza, M. C. M., Santos, A. C. M. C. & Delfini, ic structure in populations of the terrestrial orchid Cephalan-
L. T. 2004. Análise do padrão de distribuição espacial da thera longibracteata (Orchidaceae). American Journal of
araucária (Araucaria angustifolia) em algumas áreas no es- Botany 91: 52–57. https://doi.org/10.3732/ajb.91.1.52
tado do Paraná, utilizando a função K de Ripley. Scientia Dale, M. R. T. 1999. Spatial pattern analysis in plant ecol-
Forestalis 66: 38–45. ogy. Cambridge University Press, Cambridge. https://doi.
Anjos, A. M., Barberena, F. F. V. A. & Pigozzo, C. M. 2017. org/10.1017/CBO9780511612589
Biologia reprodutiva de Vanilla bahiana Hoehne (Orchida- Dearnaley, J. D. W. 2007. Further advances in orchid myc-
ceae). Orquidário 30: 67–79. orrhizal research. Mycorrhiza 17: 475–486. https://doi.
Azevedo, M. T. A., Borba, E. L., Semir, J. & Solferini, V. N. org/10.1007/s00572-007-0138-1
2007. High genetic variability in Neotropical myophilous Diário Oficial do Estado da Bahia 1987. Decreto nº 351, de
orchids. Botanical Journal of the Linnean Society 153: 22 de setembro de 1987. Retrieved April 29, 2020, from
33–40. https://doi.org/10.1111/j.1095-8339.2007.00589.x http://www.icmbio.gov.br/cepsul/images/stories/legisla-
Barberena, F. F. V. A., Baumgratz, J. F. A. & Barros, F. 2018. cao/Decretos/1987/dec_ba_351_1987_uc_criaapa_lagoas_
Ecological data for an orchid diversity hotspot show that dunasabaete_salvador_ba_altrd_dec_2540_1993_res_ce-
the subtribe Laeliinae may be endangered in the Brazilian pram_3023_2002.pdf/
Collectanea Botanica vol. 40 (enero-diciembre 2021), e001, ISSN-L: 0010-0730, https://doi.org/10.3989/collectbot.2021.v40.001Distribución espacial de Vanilla bahiana 11 Diário Oficial do Município de Salvador 2011. Decreto nº de regeneração natural em São Francisco de Paula, RS, 22.507, de 22 de dezembro de 2011. Retrieved April 7, Brasil. Floresta 42: 509–518. https://doi.org/10.5380/rf. 2020, from https://leismunicipais.com.br/a/ba/s/salvador/ v42i3.25037 decreto/2011/2250/22507/decreton-22507-2011-delimita- Krahl, A. H., Souza, G. R., Boldrini, R. F., Carneiro, B. M., -e-implanta-o-parquedas-dunas-amplia-o-parque-do-abae- Duca, C. & Azevedo-Júnior, R. R. 2010. Padrão espacial de te-e-daoutras-providencias Eltroplectris triloba (Lindl.) Pabst (Orchidaceae, Spiranthi- Diez, J. M. 2007. Hierarchical patterns of symbiotic orchid nae) em uma formação florestal de restinga no sudeste do germination linked to adult proximity and environmental Brasil. Orquidário 24: 117–123. gradients. Journal of Ecology 95: 159–170. https://doi. Krebs, C. J. 2014. Ecological methodology. Benjamin Cum- org/10.1111/j.1365-2745.2006.01194.x mings, San Francisco. Dubbern, C. A., Leal, T. S. & Pedroso-de-Moraes, C. 2013. Lima-Ribeiro, M. S. & Prado, E. C. 2006. Métodos estatísticos Distribuição espacial de Oeceoclades maculata Lindl. (Or- e estrutura espacial de populações: uma análise comparati- chidaceae) em fragmento florestal estacional semidecidual va. Acta Scientiarum. Technology 28: 219–224. https://doi. da fazenda Santa Tereza, Cordeirópolis, SP, Brasil. Nature- org/10.4025/actascitechnol.v28i2.1197 za on line 11: 29–32. Lopes, E. M., Linhares, R. G., Pires, L. O., Castro, R. N., Sou- Fajardo, C. G., Costa, R. A., Vieira, F. A. & Molina, W. F. 2015. za, G. H. M. F., Koblitz, M. G. B., Cameron, L. C. & Mac- Distribuição espacial de Cattleya granulosa: uma orquídea edo, A. F. 2019. Vanilla bahiana, a contribution from the ameaçada de extinção. Floresta e Ambiente 22: 164–170. Atlantic Forest biodiversity for the production of vanilla: A https://doi.org/10.1590/2179-8087.073714 proteomic approach through high-definition nanoLC/MS. Ferreira, A. W. C., Oliveira, M. S., Silva, E. O., Campos, Food Research International 120: 148–156. https://doi. D. S., Pansarin, E. R. & Guarçoni, E. A. E. 2017. Vanil- org/10.1016/j.foodres.2019.02.038 la bahiana Hoehne and Vanilla pompona Schiede (Or- McCormick, M. K. & Jacquemyn, H. 2013. What constrains chidaceae, Vanilloideae): two new records from Maran- the distribution of orchid populations? New Phytologist hão state, Brazil. Check List 13: 1131–1137. https://doi. 202: 392–400. https://doi.org/10.1111/nph.12639 org/10.15560/13.6.1131 Moratelli, E. M., Costa, M. D., Lovato, P. E., Santos, M. & Feuerherdt, L., Petit, S. & Jusaitis, M. 2005. Distribution of Paulilo, M. T. S. 2007. Efeito da disponibilidade de água mycorrhizal fungus associated with the endangered pink- e de luz na colonização micorrízica e no crescimento de lipped spider orchid (Arachnorchis (syn. Caladenia) behrii) Tabebuia avellanedae Lorentz ex Griseb. (Bignoniace- at Warren Conservation Park in South Australia. New Zea- ae). Revista Árvore 31: 555–566. https://doi.org/10.1590/ land Journal of Botany 43: 367–371. https://doi.org/10.108 S0100-67622007000300021 0/0028825X.2005.9512961 MMA [Ministério do Meio Ambiente do Brasil] 2012. Resolu- Flora do Brasil 2020. Vanilla bahiana. Jardim Botânico do Rio ção Conama nº 437, de 30 de dezembro de 2011. Retrieved de Janeiro, Rio de Janeiro. Retrieved April 29, 2020, from April 30, 2020, from http://www.mma.gov.br/port/conama/ http://floradobrasil.jbrj.gov.br/reflora/floradobrasil/FB12349 res/res12/Resol437.pdf Gowland, K. M., Wood, J., Clements, M. A. & Nicotra, A. Murren, C. J. & Ellison, A. M. 1998. Seed dispersal character- B. 2011. Significant phorophyte (substrate) bias is not ex- istics of Brassavola nodosa (Orchidaceae). American Jour- plained by fitness benefits in three epiphytic orchid spe- nal of Botany 85: 675–680. https://doi.org/10.2307/2446537 cies. American Journal of Botany 98: 197–206. https://doi. Nascimento, T. A., Furtado, M. S. C., Pereira, W. C. & Barber- org/10.3732/ajb.1000241 ena, F. F. V. A. 2019. Vanilla bahiana Hoehne (Orchidace- Hay, J. D., Bizerril, M. X., Calouro, A. M., Costa, E. M. N., ae): studies on fruit development and new perspectives into Ferreira, A. A., Gastal, M. L. A., Goes Junior, C. D., Man- crop improvement for the Vanilla planifolia group. Biota zan, D. J., Martins, C. R., Monteiro, J. M. G., Oliveira, S. Neotropica 19: e20180696. https://doi.org/10.1590/1676- A., Rodrigues, M. C. M., Seyffarth, J. A. S. & Walter, B. 0611-bn-2018-0696 M. T. 2000. Comparação do padrão da distribuição espacial Oksanen, J., Blanchet, F. G., Friendly, M., Kindt, R., Legendre, em escalas diferentes de espécies nativas do cerrado, em P., McGlinn, D., Minchin, P. R., O’Hara, R. B., Simpson, G. Brasília, DF. Revista Brasileira de Botânica 23: 341–347. L., Solymos, P., Stevens, M. H. H., Szoecs, E. & Wagner, https://doi.org/10.1590/S0100-84042000000300008 H. 2019. Vegan: Community Ecology Package. R package Householder, E., Janovec, J., Mozambite, A. B., Maceda, J. H., version 2.5-6. Retrieved November 3, 2020, from https:// Wells, J. & Valega, R. 2010. Diversity, natural history, and cran.r-project.org/web/packages/vegan/index.html conservation of Vanilla (Orchidaceae) in Amazonian wet- Osorio-Gil, E. M., Forero-Montaña, J. & Otero, J. T. 2008. lands of Madre de Dios, Peru. Botanical Research Institute Variation in mycorrhizal infection of the epiphytic orchid of Texas 4: 227–243. Ionopsis utriculariodes (Orchidiaceae) on different substra- Hutchings, M. J. 1997. The structure of plant populations. ta. Caribbean Journal of Science 44: 130–132. https://doi. In: Crawley, M. J. (Ed.), Plant ecology. Blackwell org/10.18475/cjos.v44i1.a15 Scientific Publications, Oxford: 325–358. https://doi. Otero, J. T., Aragón, S. & Ackerman, J. D. 2007a. Site vari- org/10.1002/9781444313642.ch11 ation in spatial aggregation and phorophyte preference in Jacquemyn, H., Brys, R., Vandepitte, K., Honnay, O., Roldán- Psychilis monensis (Orchidaceae). Biotropica 39: 227–231. Ruiz, I. & Wiegand, T. 2007. A spatially explicit analysis https://doi.org/10.1111/j.1744-7429.2006.00258.x of seedling recruitment in the terrestrial orchid Orchis Otero, J. T., Flanagan, N. S., Herre, E. A., Ackerman, J. D. & purpurea. New Phytologist 176: 448–459. https://doi. Bayman, P. 2007b. Widespread mycorrhizal specificity cor- org/10.1111/j.1469-8137.2007.02179.x relates to mycorrhizal function in the Neotropical, epiphyt- Kanieski, M. R., Longhi, S. L., Narvaes, I. S., Soares, P. R. C., ic orchid Ionopsis utricularioides (Orchidaceae). American Longhi-Santos, T. & Callegaro, R. M. 2012. Diversidade Journal of Botany 94: 1944–1950. https://doi.org/10.3732/ e padrões de distribuição espacial de espécies no estágio ajb.94.12.1944 Collectanea Botanica vol. 40 (enero-diciembre 2021), e001, ISSN-L: 0010-0730, https://doi.org/10.3989/collectbot.2021.v40.001
12 FELIPE FAJARDO VILLELA ANTOLIN BARBERENA, ESTÍBALIZ LÓPEZ HERMOSO & MARIA APARECIDA JOSÉ DE OLIVEIRA Pandey, M., Sharma, J., Donald, L. T. & Yadon, V. L. 2013. ex Miller with a characterization of their ITS region of the A narrowly endemic photosynthetic orchid is non-specif- nuclear ribosomal DNA. Lankesteriana 9: 285–354. ic in its mycorrhizal associations. Molecular Ecology 22: Sousa, D. G. & Cunha, H. F. 2018. Population structure, spatial 2341–2354. https://doi.org/10.1111/mec.12249 distribution and phenology of Anacardium humile A. St.- Pedroso-de-Moraes, C. & Souza-Leal, T. 2015. Habenaria repens: Hil. (Anacardiaceae) in cerrado stricto sensu. Hoehnea 45: distribuição espacial e correlação com fatores abióticos de uma 450–467. https://doi.org/10.1590/2236-8906-81/2017 orquídea emergente invasora em fragmento mesófilo brasileiro. Souza, G. R., Krahl, A. H. & Silva, A. G. 2012. Population Revista em Agronegócio e Meio Ambiente 8: 111–136. https:// structure and spatial pattern of Catasetum discolor (Lindl.) doi.org/10.17765/2176-9168.2015v8nEd.esp.p111-136 Lindl. (Orchidaceae) in a sandy coastal plain southeastern Peixoto, A. L. & Maia, L. C. 2013. Manual de procedimentos Brazil. Natureza on line 10: 105–110. para herbários. Editora Universitária UFPE, Recife. Souza-Leal, T. & Pedroso-de-Moraes, C. 2014. Fenologia Perkins, A. J. & McGee, P. A. 1995. Distribution of the orchid reprodutiva e distribuição espacial de Oeceoclades ma- mycorrhizal fungus, Rhizoctonia solani, in relation to its culata (Lindl.) Lindl. (Orchidaceae) em Cerrado do mu- host, Pterostylis acuminata, in the field. Australian Journal nicípio de Mogi Guaçu, São Paulo, Brasil. Iheringia 69: of Botany 43: 565–575. https://doi.org/10.1071/BT9950565 405–416. Phillips, R. D., Brown, A. P., Dixon, K. W. & Hopper, S. D. Teixeira, N. D. A., Marimon, B. S., Elias, F. & Marimon-Ju- 2010. Orchid biogeography and factors associated with nior, B. H. 2019. Padrões espaciais de samambaias em flo- rarity in a biodiversity hotspot, the Southwest Australian resta estacional perenifólia na transição Amazônia-Cerrado. Floristic Region. Journal of Biogeography 38: 487–501. Rodriguésia 70: e02572016. https://doi.org/10.1590/2175- https://doi.org/10.1111/j.1365-2699.2010.02413.x 7860201970011 Piccoli, M. C. A., Souza-Leal, T. de & Pedroso-de-Moraes, C. Tremblay, R. L., Meléndez-Ackerman, E. & Kapan, D. 2006. 2014. Distribuição espacial de Sacoila lanceolata (Aubl.) Do epiphytic orchids behave as metapopulations? Evidence Garay (Orchidaceae) em fragmento mesófilo de Pirassu- from colonization, extinction rates and asynchronous pop- nunga, São Paulo, Brasil. Nucleus 11: 115–130. https://doi. ulation dynamics. Biological Conservation 129: 70–81. org/10.3738/1982.2278.937 https://doi.org/10.1016/j.biocon.2005.11.017 Porras-Alfaro, A. & Bayman, P. 2007. Mycorrhizal fungi of Viana, B. F. & Kleinert, A. M. P. 2005. A community of flow- Vanilla: diversity, specificity and effects on seed germina- er-visiting bees (Hymenoptera: Apoidea) in the coastal sand tion and plant growth. Mycologia 99: 510–525. https://doi. dunes of Northeastern Brazil. Biota Neotropica 5: 1–13. org/10.3852/mycologia.99.4.510 https://doi.org/10.1590/S1676-06032005000300005 Quenzer, F. C. L. & Pedroso-de-Moraes, C. 2014. Distribuição Waterman, R. J. & Bidartondo, M. I. 2008. Deception above, espacial de Oeceoclades maculata Lindl. em fragmento flo- deception below: linking pollination and mycorrhizal biol- restal estacional semidecidual do “Sítio João XXIII”, Piras- ogy of orchids. Journal of Experimental Botany 59: 1085– sununga, SP, Brasil. Natureza on line 12: 160–163. 1096. https://doi.org/10.1093/jxb/erm366 Rahal, M. H., Souza-Leal, T. & Pedroso-de-Moraes, C. 2015. Watteyn, C., Fremout, T., Karremans, A. P., Pillco-Huarcaya, Ecologia e distribuição espacial de orquídeas terrícolas em R., Azofeifa-Bolaños, J. B., Reubens, B. & Muys, B. 2020. floresta estacional semidecidual do município de Araras, Vanilla distribution modeling for conservation and sus- São Paulo, Brasil. Iheringia 70: 217–228. tainable cultivation in a joint land sparing/sharing con- Soto-Arenas, M. A. & Dressler, R. L. 2010. A revision of the cept. Agroecosystems 11: e03056. https://doi.org/10.1002/ Mexican and Central American species of Vanilla Plumier ecs2.3056 Collectanea Botanica vol. 40 (enero-diciembre 2021), e001, ISSN-L: 0010-0730, https://doi.org/10.3989/collectbot.2021.v40.001
También puede leer

















































