UN ANTICUERPO MONOCLONAL HUMANO QUE BLOQUEA LA INFECCIÓN POR SARS-COV-2
←
→
Transcripción del contenido de la página
Si su navegador no muestra la página correctamente, lea el contenido de la página a continuación
1
UN ANTICUERPO MONOCLONAL HUMANO QUE
BLOQUEA LA INFECCIÓN POR SARS-COV-2
Wang, C., Li, W., Drabek, D. et al. A human monoclonal antibody blocking SARS-CoV-2 infection. Nat
Commun 11, 2251 (2020). https://doi.org/10.1038/s41467-020-16256-y
INTRODUCCIÓN
El síndrome respiratorio agudo severo por coronavirus 2 (SARS-CoV-2) es el agente etiológico de la
enfermedad inducida por coronavirus 19 (COVID-19) que surgió en China a fines de 2019 y causó una
pandemia (1). Hasta el 19 de abril de 2020, se habían notificado 2.241.778 casos en todo el mundo, de
los cuales 152.551 (6,8%) sucumbieron a la infección (2).
El SARS-CoV-2 pertenece al subgénero Sarbecovirus (género Betacoronavirus, familia Coronaviridae)
(3) junto con el SARS-CoV que surgió en 2002 causando aproximadamente 8000 infecciones con una
letalidad del 10%. Ambos virus cruzaron las barreras de especies de un reservorio animal, y pueden
causar una enfermedad respiratoria potencialmente mortal en humanos. Actualmente, no existen
terapias dirigidas aprobadas para COVID-19. Los anticuerpos monoclonales dirigidos a sitios
vulnerables en las proteínas de la superficie viral, se reconocen cada vez más, como una clase
prometedora de medicamentos contra enfermedades infecciosas y han demostrado eficacia
terapéutica para varios virus (4,5).
Los anticuerpos neutralizantes contra coronavirus se dirigen principalmente a las glicoproteínas
triméricas de la espiga (S), que se encuentran en la superficie viral y que median la entrada en las
células hospedadoras. La proteína S tiene dos subunidades funcionales que median la unión celular (la
subunidad S1, que presenta cuatro dominios centrales S1A a S1D) y la fusión de la membrana viral y
celular (la subunidad S2). Los anticuerpos neutralizantes potentes, a menudo se dirigen al sitio de
interacción del receptor en S1, deshabilitando las interacciones del receptor (6,7,8,9,10,11). Las
proteínas de pico de SARS-CoV-2 (SARS2-S; 1273 residuos, cepa Wuhan-Hu-1) y SARS-CoV (SARS-S,
1255 residuos, cepa Urbani) son 77.5% idénticas por la secuencia de aminoácidos primaria, son
estructuralmente muy similares (12,13,14,15) y comúnmente se unen a través de su dominio S1B, al
receptor de la enzima covertidora de angiotensina humana 2 (ACE2) (1,16). Se sabe que la interacción
del receptor desencadena cambios conformacionales irreversibles en las proteínas de la espiga del
coronavirus, que permiten la fusión de la membrana (17).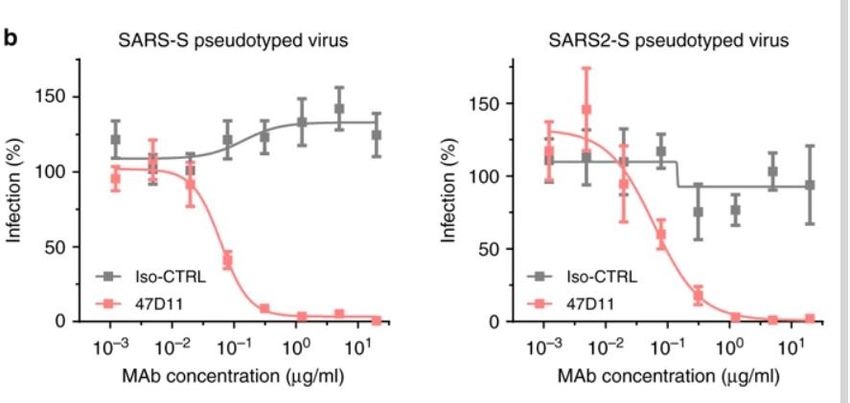
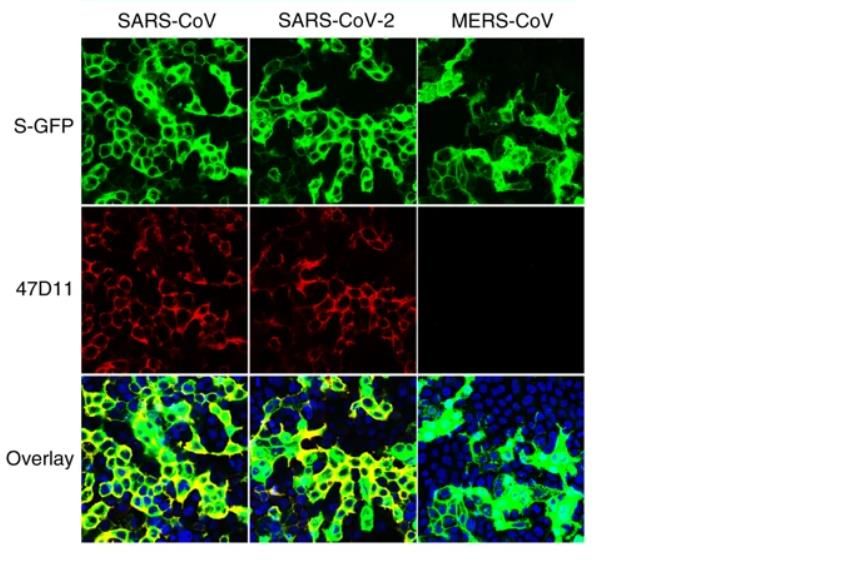
2 RESULTADOS IDENTIFICACIÓN DE ANTICUERPOS REACTIVOS CONTRA EL SARS-COV-2 Con el fin de identificar los anticuerpos neutralizantes contra el SARS-CoV-2, se evaluó la reactividad ELISA (cruzada) de los sobrenadantes que contienen anticuerpos de una colección de 51 hibridomas de SARS-S derivados de ratones transgénicos H2L2 inmunizados que codifican inmunoglobulinas quiméricas con dominios variables de cadenas pesadas y livianas, de inmunoglobulinas humanas y regiones constantes de rata (Tabla complementaria 1). Cuatro de 51 sobrenadantes de hibridoma de SARS-S anticuerpos reactividad cruzada ELISA con la subunidad SARS2-S1 (residuos S 1-681; Tabla complementaria 1), de los cuales uno (47D11) exhibió actividad de neutralización cruzada de SARS-S y SARS2 - S infección por VSV seudotipado. El anticuerpo quimérico 47D11 H2L2 se volvió a formatear en una inmunoglobulina completamente humana, mediante la clonación de las regiones variables de cadena pesada y ligera humana, en un esqueleto de isotipo IgG1 humano. El anticuerpo 47D11 humano expresado recombinantemente, se usa para una caracterización adicional. PROPIEDADES ANTIVIRALES Y BIOQUÍMICAS DEL ANTICUERPO MONOCLONAL HUMANO 47D11 El anticuerpo 47D11 humano, se une a las células que expresan las proteínas de la espiga de longitud completa de SARS-CoV y SARS-CoV-2 (Figura 1a). Se encontró que el anticuerpo 47D11 inhibía de manera potente la infección de las células VeroE6 con VSV pseudotipado SARS-S y SARS2-S con valores de CI50 de 0.061 y 0.061 μg / ml (Figura 1b), respectivamente. La infección auténtica de las células VeroE6 con SARS-CoV y SARS-CoV-2 se neutralizó con valores de CI50 de 0.19 y 0.57 μg / ml (Figura 1c). Se demostró que el uso de ELISA 47D11 apunta al dominio de unión al receptor S1B (RBD) de SARS- S y SARS2-S. el 47D11 se unió al S1B de ambos virus con afinidades similares a las mostradas por los valores de concentración efectiva media máxima (CE50) basados en ELISA (0.02 y 0.03 μg / ml, respectivamente; Figura 2a). La afinidad de unión, basada en un ELISA de 47D11 para el ectodominio de la proteína S (Secto) de SARS-CoV, fue mayor en relación con la de SARS-CoV-2 (valores de CE50: 0,018 y 0,15 μg / ml, respectivamente), a pesar del recubrimiento de antígeno equimolar (Figura complementaria 1) Congruente con las reactividades por ELISA, la medición de la cinética de unión de 47D11 por interferometría de biocapa mostró que 47D11 une SARS-Secto con mayor afinidad (constante de disociación de equilibrio [KD]: 0.745 nM) en relación con SARS2-Secto (KD 10.8 nM), mientras que la afinidad para SARS-S1B y SARS2-S1B estuvo en un rango similar (16.1 y 9.6 nM, respectivamente, Figura 2 complementaria). Esta diferencia puede originarse de diferencias en la accesibilidad del epitope en SARS-S versus SARS2-S, ya que el dominio B puede adoptar una conformación cerrada y abierta en el homotrímero de pico con conformación de prefusión (12,13). Sorprendentemente, la unión del 47D11 al SARS-S1B y SARS2-S1B no compitió con la unión de S1B al receptor ACE2 expresado en la superficie celular como se muestra por citometría de flujo (Figura 2b; Figura Suplementaria 3) ni con la unión de Secto y S1B a ACE2 soluble en un ensayo basado en fase sólida (Figura 4 suplementaria), mientras que dos anticuerpos específicos contra SARS-S1 35F4 y 43C6 que neutralizan la infección por VSV pseudotipada SARS-S (pero no SARS2-S) (Figura complementaria 5) bloquean la unión de SARS-Secto y SARS-S1B a ACE2. Usando un ensayo de fusión célula-célula desencadenado por tripsina, se demostró que el 47D11 perjudica la formación de sincitios mediada por SARS-S y SARS2-S (Figura complementaria 6). Nuestros datos muestran que el 47D11 neutraliza el SARS-CoV y el SARS-CoV-2 a través de un mecanismo aún desconocido que es diferente de la interferencia de unión al receptor. Se han informado mecanismos alternativos de neutralización del coronavirus por anticuerpos dirigidos a RBD, incluida la inactivación

3
de la espiga a través de la desestabilización inducida por anticuerpos de su estructura de prefusión
(17), que también puede aplicarse para el 47D11.
Figura1- El 47D11 neutraliza SARS-CoV y SARS-CoV-2.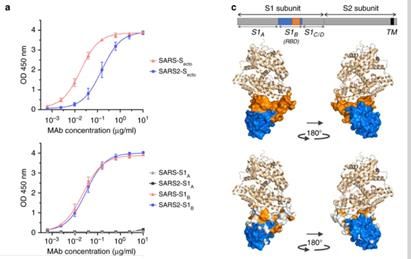
4
FIGURA 2- EL 47D11 NEUTRALIZANTE SE UNE A LA SARS S-1 Y SARS S-2
RBD SIN ELIMINAR LA INTERACCIÓN DEL RECEPTOR.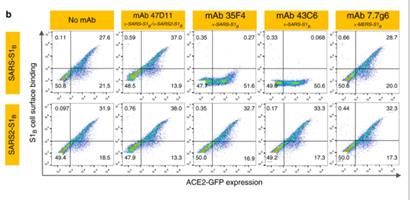
5 Curiosamente, recientemente se descubrió que el anticuerpo neutralizador de SARS-CoV CR3022 dirigido también al dominio central S1B se unía de forma cruzada con SARS-CoV-2, aunque no se informó su capacidad para neutralizar de forma cruzada la infección por SARS-CoV-218,20. La unión de S1B por el 47D11 más lejos de la interfaz de unión al receptor, explica su incapacidad para comprometer la interacción espiga-receptor y abre posibilidades para tratamientos combinados con potentes anticuerpos neutralizantes no competitivos que se dirijan al subdominio de unión al receptor. Las combinaciones de anticuerpos dirigidas a epitopes no superpuestos, pueden actuar de forma sinérgica, lo que resulta en una dosis más baja y pueden mitigar el riesgo del escape inmune (20). En conclusión, este es el primer informe de un anticuerpo monoclonal (humano) que neutraliza el SARS-CoV-2. El 47D11 se une a un epitope conservado en el pico RBD explicando su capacidad para neutralizar en forma cruzada al SARS-CoV y SARS-CoV-2, utilizando un mecanismo que es independiente de la inhibición de unión al receptor. Este anticuerpo será útil para el desarrollo de pruebas de detección de antígeno y ensayos serológicos dirigidos al SARS-CoV-2. Los anticuerpos neutralizantes, pueden alterar el curso de la infección en el hospedador infectado que respalda la eliminación del virus o proteger a un hospedador no infectado que está expuesto al virus (4). Por lo tanto, este anticuerpo, ya sea solo o en combinación, ofrece el potencial para prevenir y tratar COVID- 19, y posiblemente también otras enfermedades emergentes en humanos causadas por virus del subgénero Sarbecovirus. ANÁLISIS POR TÉCNICA ELISA DE UNIÓN DE ANTICUERPOS A ANTÍGENOS (ESPIGA) DEL COV Las placas NUNC Maxisorp (Thermo Scientific) se recubrieron con cantidades de antígeno equimolar a 4 ° C durante la noche. Las placas se lavaron tres veces con PBS que contenía Tween-20 al 0,05% y se bloquearon con albúmina de suero bovino al 3% (Bio-Connect) en PBS que contenía Tween-20 al 0,1% a temperatura ambiente durante 2 horas. Se añadieron diluciones en serie cuádruples de mAbs que comenzaban a 10 µg / ml (diluido en tampón de bloqueo) y las placas se incubaron durante 1 hora a temperatura ambiente. Las placas se lavaron tres veces y se incubaron con un anticuerpo secundario antihumano de cabra conjugado con peroxidasa de rábano picante (HRP) (ITK Southern Biotech) diluido 1: 2000 en tampón de bloqueo durante 1 hora a temperatura ambiente. Se usó un anticuerpo anti-StrepMAb conjugado con HRP (IBA, Catálogo # 2-1509-001) para corroborar el recubrimiento equimolar de los antígenos de la espiga marcados. La actividad de HRP se midió a 450 nanómetros usando como sustrato tetrametilbencidina (BioFX) y un lector de placas de ELISA (EL-808, Biotek). Los valores de unión de la concentración efectiva media máxima (CE50) se calcularon mediante análisis de regresión no lineal en las curvas de unión usando GraphPad Prism (versión 8). BIBLIOGRAFÍA 1. Zhou, P. et al. A pneumonia outbreak associated with a new coronavirus of probable bat origin. Nature 579, 1–4 (2020). 2. World Health Organization. https://www.who.int/docs/default-source/ coronaviruse/situation-reports/20200419-sitrep-90-covid- 19.pdf? sfvrsn=551d47fd_4. 3. Coronaviridae Study Group of the International Committee on Taxonomy of Viruses. The species severe acute respiratory syndrome-related coronavirus: classifying 2019-nCoV and naming it SARS-CoV-2. Nat. Microbiol. 5, 536–544 (2020). 4. Prabakaran, P. et al. Potent human monoclonal antibodies against SARS CoV, Nipah and Hendra viruses. Expert Opin. Biol. Ther 9, 355–368 (2009). 5. Saphire, E. O., Schendel, S. L., Gunn, B. M., Milligan, J. C. & Alter, G. Antibody-mediated protection against Ebola virus. Nat. Immunol. 19, 1169–1178 (2018). 6. Reguera, J. et al. Structural bases of coronavirus attachment to host aminopeptidase N and its inhibition by neutralizing antibodies. PLoS Pathog.8, e1002859 (2012).
6 7. Yu, X. et al. Structural basis for the neutralization of MERS-CoV by a human monoclonal antibody MERS-27. Sci. Rep. 5, 13133 (2015). 8. Prabakaran, P. et al. Structure of severe acute respiratory síndrome coronavirus receptor-binding domain complexed with neutralizing antibody. J. Biol. Chem. 281, 15829–15836 (2006). 9. Hwang, W. C. et al. Structural basis of neutralization by a human anti-severe acute respiratory syndrome spike protein antibody, 80R. J. Biol. Chem. 281, 34610–34616 (2006). 10. Rockx, B. et al. Structural basis for potent cross-neutralizing human monoclonal antibody protection against lethal human and zoonotic severe acute respiratory syndrome coronavirus challenge. J. Virol. 82, 3220–3235 (2008). 11. Widjaja, I. et al. Towards a solution to MERS: protective human monoclonal antibodies targeting different domains and functions of the MERScoronavirus spike glycoprotein. Emerg. Microbes Infect. 8, 516–530 (2019). 12. Wrapp, D. et al. Cryo-EM structure of the 2019-nCoV spike in the prefusion conformation. Science 367, 1260–1263 (2020). 13. Walls, A. C. et al. Structure, function, and antigenicity of the SARS-CoV-2 spike glycoprotein. Cell 181, 281–292.e6 (2020). 14. Yuan, Y. et al. Cryo-EM structures of MERS-CoV and SARS-CoV spike glycoproteins reveal the dynamic receptor binding domains. Nat. Commun. 8, 15092 (2017). 15. Gui, M. et al. Cryo-electron microscopy structures of the SARS-CoV spike glycoprotein reveal a prerequisite conformational state for receptor binding. Cell Res. 27, 119–129 (2017). 16. Li, W. et al. Angiotensin-converting enzyme 2 is a functional receptor for the SARS coronavirus. Nature 426, 450–454 (2003). 17. Walls, A. C. et al. Unexpected receptor functional mimicry elucidates activation of coronavirus fusion. Cell 176, 1026–1039.e15 (2019). 18. Tian, X. et al. Potent binding of 2019 novel coronavirus spike protein by a SARS coronavirus-specific human monoclonal antibody. Emerg. Microbes Infect. 9, 382–385 (2020). 19. Menachery, V. D. et al. A SARS-like cluster of circulating bat coronaviruses shows potential for human emergence. Nat. Med. 21, 1508 (2015). 20. ter Meulen, J. et al. Human monoclonal antibody combination against SARS coronavirus: synergy and coverage of escape mutants. PLoS Med. 3, e237 (2006). 21. Raj, V. S. et al. Dipeptidyl peptidase 4 is a functional receptor for the emerging human coronavirus-EMC. Nature 495, 251– 254 (2013). 22. Bosch, B. J., Bartelink, W. & Rottier, P. J. Cathepsin L functionally cleaves the severe acute respiratory syndrome coronavirus class I fusion protein upstream of rather than adjacent to the fusion peptide. J. Virol. 82, 8887–8890 (2008). 23. Okba, N. M. A. et al. Sensitive and specific detection of low-level antibody responses in mild middle east respiratory syndrome coronavirus infections. Emerg. Infect. Dis. 25, 1868–1877 (2019). 24. Li, F., Li, W., Farzan, M. & Harrison, S. C. Structure of SARS coronavirus spike receptor-binding domain complexed with receptor. Science 309, 1864–1868 (2005
También puede leer

















































