Un Sustrato Neuronal Común entre Varios Anestésicos y el Sueño
←
→
Transcripción del contenido de la página
Si su navegador no muestra la página correctamente, lea el contenido de la página a continuación
SNACC Newsletter ▪ Fall 2019 Issue ▪ Spanish Translation
Fall 2019 Issue ▪ Spanish Translation
Un Sustrato Neuronal Común entre Varios
Anestésicos y el Sueño
Luping Yin1, Li-Feng Jiang-Xie1, Fan Wang1
1Departmento de Neurobiología, Duke University Medical Center
Durham, North Carolina
Antecedentes
Los anestésicos generales se han utilizado ampliamente en cirugía desde 1846, cuando William T.G. Morton
(1819-1868) realizó la primera anestesia general (GA) moderna con éxito en el Hospital General de
Massachusetts1. El descubrimiento de la anestesia general ha revolucionado los procedimientos quirúrgicos
en medicina durante las últimas décadas1,2. Cada año, millones de pacientes en todo el mundo reciben GA
para tratamientos médicos. La GA es un estado cerebral / corporal reversible y global inducido por fármacos
que se caracteriza por perdida de la conciencia, amnesia, analgesia e inmovilidad junto con la estabilidad de
las funciones fisiológicas básicas2,3. A pesar de siglos de esfuerzo científico, los mecanismos del circuito
neural por los cuales grupos de anestésicos generales de diversa estructura inducen perdida de la conciencia
permanecen confusos3-5.
El proceso de GA puede dividirse en tres períodos según los signos clínicos y los patrones del
electroencefalograma (EEG): inducción, mantenimiento y emersión. Los patrones de EEG durante el período
de mantenimiento se caracterizan por la disminución progresiva de la potencia de las oscilaciones de
frecuencia más alta (13-30 Hz) y la aparición de oscilación delta de baja frecuencia y alta amplitud (0.5-4 Hz).
Curiosamente, el aumento de la oscilación delta se asemeja a lo que ocurre durante el sueño natural de onda
lenta2,6,7. La identificación precisa de las neuronas y los circuitos neuronales compartidos entre la GA y el
sueño, debería enriquecer nuestra comprensión básica y las aplicaciones clínicas de la GA.
Blancos moleculares de la anestesia general
Page | 1
©2019 Society for Neuroscience in Anesthesiology and Critical Care.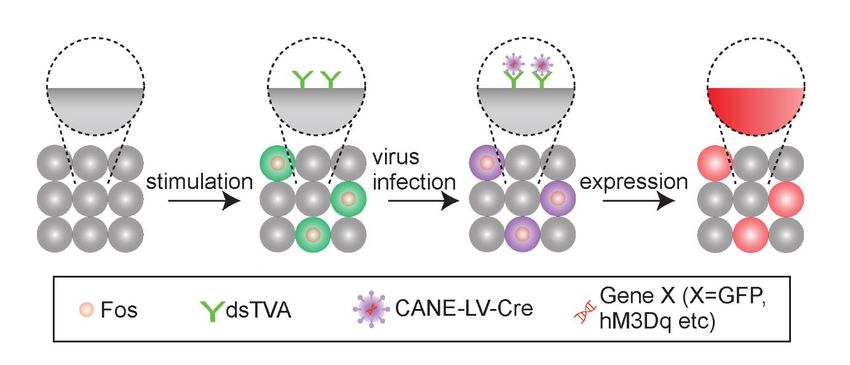
SNACC Newsletter ▪ Fall 2019 Issue ▪ Spanish Translation
Se han descubierto muchos receptores y blancos moleculares diferentes para diversos anestésicos generales.
Se cree que el estado de GA es consecuencia de la mejora de la inhibición o la supresión de la
neurotransmisión excitadora. Entre los blancos moleculares conocidos de los anestésicos, los canales iónicos
son los candidatos predominantes. Se ha demostrado que muchas drogas como el isoflurano, anestésico
gaseoso, y los anestésicos intravenosos como barbitúricos y propofol, potencian la corriente de Cl- inducida
por GABA; en dosis más altas, pueden activar directamente los receptores GABAA incluso sin anestésicos2.
Se ha demostrado que otros anestésicos no GABA, como la ketamina, reducen la transmisión sináptica
excitadora o mediada por glutamato al bloquear los receptores NMDA en la membrana postsináptica y la
dexmedetomidina, que se une a los receptores adrenérgicos alfa-2 (α2) e inhiben la liberación de
noradrenalina desde el locus coeruleus2. Los canales de sodio / potasio y otros canales iónicos como los
canales de nucleótidos cíclicos activados por hiperpolarización (HCN) también están involucrados en este
proceso. Colectivamente, estos resultados conducen a la creencia común de que distintos anestésicos
funcionan al ejercer distintos efectos inhibitorios sobre el sistema nervioso. En los últimos años, algunos
estudios han sugerido la existencia de células activadas por anestesia en varias regiones del cerebro y han
cuestionado esta justificación utilizando marcadores genéticos tempranos inmediatos c-fos o registro de corte
cerebral ex vivo 8-11, pero aún se necesita evidencia in vivo del registro directo extracelular.
Tecnología utilizada
Utilizando el primitivo gen Fos, hemos descubierto una población de neuronas ubicadas en la región
inmediatamente superior al tracto óptico: el núcleo supraóptico y sus regiones cercanas. Llamamos a esta
población distinta de neuronas "neuronas activadas por anestesia (AAN)". Nuestra hipótesis novedosa es que
las AAN juegan papeles esenciales en el proceso de la GA y también pueden estar involucrados en la
regulación del sueño. Para etiquetar y manipular las neuronas Fos + activadas para probar esta hipótesis,
utilizamos una tecnología de vanguardia llamada "CANE" para "capturar conjuntos neuronales activados"
recientemente desarrollados en nuestro laboratorio 12 (Figura 1). La tecnología CANE consta de dos partes:
(1) un ratón knock-in en el que un receptor extraño desestabilizado (dsTVA) se expresa en un patrón espacial
y temporal similar al del gen primitivo Fos (ratón FosTVA), y (2) virus de diseño pseudotipado (CANE-lentivirus
o CANE-rabia) que pueden transportar los transgenes deseados y solo infectan las neuronas que expresan el
receptor dsTVA, por lo tanto, las neuronas Fos +. Mostramos que CANE puede usarse para capturar de
manera específica y eficiente las neuronas Fos + y expresar los genes deseados en las neuronas activadas.
Por lo tanto, CANE nos proporciona una herramienta sin precedentes para lograr este objetivo con una
Page | 2
©2019 Society for Neuroscience in Anesthesiology and Critical Care.
SNACC Newsletter ▪ Fall 2019 Issue ▪ Spanish Translation
resolución temporal precisa.
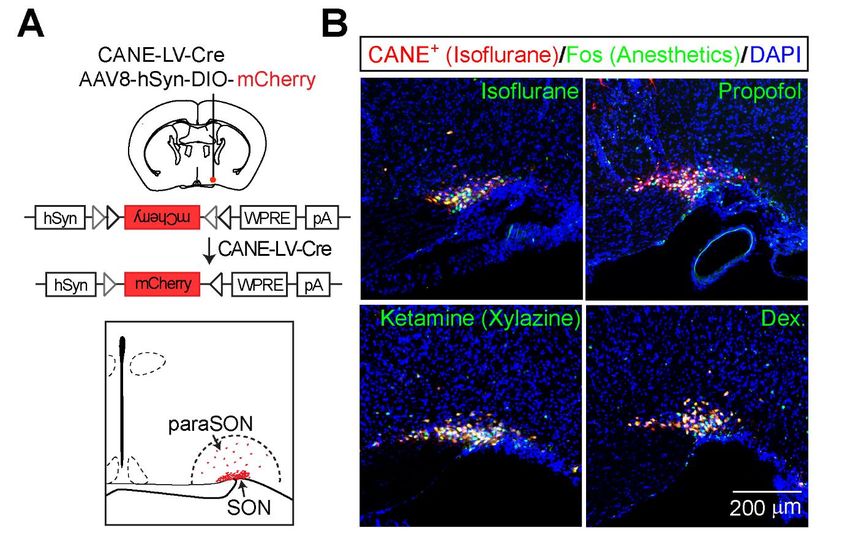
Figura 1 Representación esquemática de la tecnología CANE
Principales hallazgos de nuestro estudio
En nuestro reciente trabajo publicado13, mostramos que la mayoría de las AAN se encontraban dentro de un
pequeño núcleo del cerebro, llamado núcleo supraóptico (SON) (Figura 2A), que se conoce como el centro de
las células neuroendocrinas que liberan hormonas para mantener la homeostasis y estas AAN expresan
consistentemente múltiples neuropéptidos y hormonas, incluyendo vasopresina, dinorfina y galanina.
Utilizamos el registro electrofisiológico in vivo para revelar que los subconjuntos de neuronas dentro y cerca
del SON aumentaron significativamente su activación in vivo en respuesta a la anestesia con isoflurano.
Después de usar CANE para capturar y etiquetar AAN, demostramos que las AAN pueden activarse mediante
múltiples anestésicos generales distintos ampliamente utilizados en ensayos clínicos, que incluyen isoflurano,
propofol, ketamina / xilazina y dexmedetomidina, usando tinción con Fos (Figura 2B) y cortes
electrofisiológicos.
Page | 3
©2019 Society for Neuroscience in Anesthesiology and Critical Care.SNACC Newsletter ▪ Fall 2019 Issue ▪ Spanish Translation
Figura 2 A Conglomerado Común de Neuronas Activadas por Diferentes Anestésicos
(A) Arriba, construcción viral y sitio de inyección en ratones FosTVA. Abajo, ilustración del SON y paraSON. (B)
Imágenes representativas de neuronas activadas por isoflurano capturadas por CANE (rojo) y neuronas Fos +
(verde) inducidas por la reexposición a isoflurano, o a Propofol, Ketamina (más xilazina) o dexmedetomidina
(Dex).
Queríamos manipular selectivamente estas AAN y observamos la consecuencia en los comportamientos de
los animales. Para lograr este objetivo, empleamos la tecnología CANE para expresar específicamente
actuadores quimiogenéticos (hM3Dq-DREADD) u optogenéticos (Channelrhodopsin, ChR2) en estas AAN, o
proteínas fluorescentes de control. Sorprendentemente, una vez que activamos las AAN, usando el ligando
artificial CNO que funciona en el receptor hM3Dq-DREADD o con un pulso de luz azul para estimular la
expresión de ChR2 en las AAN, los ratones dejaron de moverse. Las grabaciones concomitantes de EEG
mostraron que su estado cerebral cambió de vigilia a un estado de sueño profundo caracterizado por una
acentuada oscilación de onda lenta. Incluso una breve activación de las AAN es suficiente para dar como
resultado un sueño prolongado, consistente con el hecho de que los neuropéptidos liberados por las mismas
probablemente tengan una señalización más duradera después de que se apagó el pulso de luz. Por otro
lado, luego de la ablación específica de estas AAN, los ratones reducen el tiempo total de sueño, tienen un
Page | 4
©2019 Society for Neuroscience in Anesthesiology and Critical Care.SNACC Newsletter ▪ Fall 2019 Issue ▪ Spanish Translation
sueño fragmentado e incluso si duermen, su cerebro muestra una oscilación lenta disminuida. Algunos
ratones, post-ablación de las AAN, murieron por falta de sueño.
Además, también investigamos el papel funcional de las AAN durante la GA. Al expresar selectivamente ChR2
para activar estas células, o al expresar una bomba de protones activada por luz, eArch3.0 para apagar las
células con luz, manipulamos bidireccionalmente la actividad de las AAN durante la anestesia. La
fotoactivación de las AAN no alteró la inducción de la GA, pero retrasó significativamente la aparición de la
misma. En otras palabras, los animales permanecerían más tiempo bajo GA una vez que activemos las AAN.
Por el contrario, la inhibición de las AAN no alteró la inducción de GA, pero acortó significativamente la
duración de la misma. Estos resultados subrayan aún más la importancia de la señalización peptídica
(hormonal) lenta pero duradera de las AAN para mantener el estado de GA. En resumen, las AAN tienen un
papel crucial en el mantenimiento, pero no en la inducción de la perdida de la conciencia inducida por la GA.
Contribución al Campo de la Anestesia General y del Sueno
Las AAN descubiertas aquí son activadas por múltiples anestésicos diferentes y pueden representar un
sustrato compartido entre la GA y el sueño. En ratones de comportamiento libre, la activación optogenética14 y
quimiogenética15 de las AAN fue suficiente para potenciar fuertemente el sueño de onda lenta (SWS) y GA. Es
importante destacar que la ablación condicional de las AAN resultó en una disminución significativa de la
potencia de onda lenta y la pérdida tanto del SWS como del sueño de movimiento ocular rápido (REM),
mientras que la inhibición aguda de las AAN acortó la duración de la GA. Juntos, nuestros resultados
revelaron una función previamente no reconocida de las células neuroendocrinas SON (principalmente células
productoras de vasopresina / dinorfina / galanina en SON), que son conocidas por su papel de liberar
hormonas, en la regulación de la GA y del sueño natural.
Significancia
Las AAN representan un sustrato neuronal común para la GA y el sueño, y funcionan para mantener o
prolongar la perdida de la conciencia y el sueño. El descubrimiento más sorprendente es que la mayoría de
las AAN son células neuroendocrinas liberadoras de péptidos en el SON. Resulta que estas células no solo
liberan hormonas peptídicas en la circulación a través de sus proyecciones a la hipófisis posterior, sino que
también liberan grandes cantidades de péptidos a través de la liberación dendrítica masiva en el hipotálamo y
los fluidos cerebroespinales. Especulamos que la señalización de neuropéptidos puede durar más tiempo y a
través del líquido cefalorraquídeo diseminado por todo el cerebro, y por lo tanto puede ser ideal para mantener
el estado cerebral global en un estado de pérdida de la conciencia.
Page | 5
©2019 Society for Neuroscience in Anesthesiology and Critical Care.SNACC Newsletter ▪ Fall 2019 Issue ▪ Spanish Translation
Agradecimientos
Este trabajo es financiado con fondos del NIH DP1MH103908 para F.W., Brain Research Foundation SIA para
F.W., W. M. Keck Foundation para F.W. and K.D, y el Human Frontier Science Program (LT000038/2018-L)
para L.Y.
Referencias
1. Robinson, D. H. & Toledo, A. H. Historical development of modern anesthesia. J Invest Surg 25, 141-
149, doi:10.3109/08941939.2012.690328 (2012).
2. Franks, N. P. General anaesthesia: from molecular targets to neuronal pathways of sleep and arousal.
Nat Rev Neurosci 9, 370-386, doi:10.1038/nrn2372 (2008).
3. Brown, E. N., Lydic, R. & Schiff, N. D. General anesthesia, sleep, and coma. N Engl J Med 363, 2638-
2650, doi:10.1056/NEJMra0808281 (2010).
4. Alkire, M. T., Hudetz, A. G. & Tononi, G. Consciousness and anesthesia. Science 322, 876-880,
doi:10.1126/science.1149213 (2008).
5. Koch, C., Massimini, M., Boly, M. & Tononi, G. Neural correlates of consciousness: progress and
problems. Nat Rev Neurosci 17, 307-321, doi:10.1038/nrn.2016.22 (2016).
6. Akeju, O. & Brown, E. N. Neural oscillations demonstrate that general anesthesia and sedative states
are neurophysiologically distinct from sleep. Curr Opin Neurobiol 44, 178-185,
doi:10.1016/j.conb.2017.04.011 (2017).
7. Rudolph, U. & Antkowiak, B. Molecular and neuronal substrates for general anaesthetics. Nat Rev
Neurosci 5, 709-720, doi:10.1038/nrn1496 (2004).
8. Gelegen, C. et al. Excitatory Pathways from the Lateral Habenula Enable Propofol-Induced Sedation.
Curr Biol 28, 580-587 e585, doi:10.1016/j.cub.2017.12.050 (2018).
9. Lu, J. et al. Role of endogenous sleep-wake and analgesic systems in anesthesia. J Comp Neurol 508,
648-662, doi:10.1002/cne.21685 (2008).
10. Moore, J. T. et al. Direct activation of sleep-promoting VLPO neurons by volatile anesthetics contributes
to anesthetic hypnosis. Curr Biol 22, 2008-2016, doi:10.1016/j.cub.2012.08.042 (2012).
11. Zhang, Z. et al. Neuronal ensembles sufficient for recovery sleep and the sedative actions of alpha2
adrenergic agonists. Nat Neurosci 18, 553-561, doi:10.1038/nn.3957 (2015).
Page | 6
©2019 Society for Neuroscience in Anesthesiology and Critical Care.SNACC Newsletter ▪ Fall 2019 Issue ▪ Spanish Translation
12. Sakurai, K. et al. Capturing and Manipulating Activated Neuronal Ensembles with CANE Delineates a
Hypothalamic Social-Fear Circuit. Neuron 92, 739-753, doi:10.1016/j.neuron.2016.10.015 (2016).
13. Jiang-Xie, L.-F. et al. A Common Neuroendocrine Substrate for Diverse General Anesthetics and
Sleep. Neuron 102, 1053-1065.e1054, doi:https://doi.org/10.1016/j.neuron.2019.03.033 (2019).
14. Boyden, E. S., Zhang, F., Bamberg, E., Nagel, G. & Deisseroth, K. Millisecond-timescale, genetically
targeted optical control of neural activity. Nat Neurosci 8, 1263-1268, doi:10.1038/nn1525 (2005).
15. Roth, B. L. DREADDs for Neuroscientists. Neuron 89, 683-694, doi:10.1016/j.neuron.2016.01.040
(2016).
Page | 7
©2019 Society for Neuroscience in Anesthesiology and Critical Care.También puede leer

















































