CARACTERIZACIÓN DE LA CAPACIDAD DISPERSIVA DE HIERBAS ANUALES DEL OLIVAR. UNA APROXIMACIÓN MEDIANTE MODELOS MECANICISTAS Y TÚNEL DE - TAUJA
←
→
Transcripción del contenido de la página
Si su navegador no muestra la página correctamente, lea el contenido de la página a continuación
Centro de Estudios de Postgrado
UNIVERSIDAD DE JAÉN
Centro de Estudios de Postgrado
Trabajo Fin de Máster
CARACTERIZACIÓN DE LA
CAPACIDAD DISPERSIVA DE
HIERBAS ANUALES DEL
OLIVAR. UNA APROXIMACIÓN
MEDIANTE MODELOS
MECANICISTAS Y TÚNEL DE
VIENTO.
Autor: Víctor Valenzuela Polo
Jaén, octubre del 2021
UNIVERSIDAD DE JAÉN
Universidad de Jaén
Centro de Estudios de Postgrado
MÁSTER EN ANÁLISIS, CONSERVACIÓN Y RESTAURACIÓN DE COMPONENTES
FÍSICOS Y BIÓTICOS DE LOS HÁBITAT
CARACTERIZACIÓN DE LA
CAPACIDAD DISPERSIVA DE HIERBAS
ANUALES DEL OLIVAR. UNA
APROXIMACIÓN MEDIANTE MODELOS
MECANICISTAS Y TÚNEL DE VIENTO.
Tutores:
Mª Rocío Bolaños Jiménez Antonio José Manzaneda Ávila
BOLAÑOS Firmado
Antonio
Firmado
digitalmente por digitalmente
JIMENEZ BOLAÑOS JIMENEZ por Antonio
MARIA MARIA ROCIO -
77329190Q
Manzan Manzaneda
Fecha:
ROCIO -
77329190Q
Fecha: 2021.10.07
12:41:06 +02'00' eda 2021.10.07
12:55:25 +02'00'

Resumen
Existe una gran cantidad de literatura científica acerca de los procesos dispersivos
anemócoros con el fin de entenderlos y relacionarlos con otros procesos ecológicos como la
persistencia de las poblaciones y comunidades, procesos evolutivos, colonizaciones, etc.
Sin embargo, los trabajos y estudios de parametrización y cuantificación de estos
procesos de dispersión no son todavía muy abundantes. Este trabajo se centrará en la
dispersión a través del vector del viento, es decir, la dispersión anemócora e intentará
realizar una caracterización, medición de las variables de peso en el proceso y predicción de
la capacidad dispersiva de algunas especies herbáceas anuales del agroecosistema del
olivar. Esto se llevará a cabo mediante una aproximación a través de modelos mecanicistas
como el modelo de WALD y ecuaciones balísticas de dispersión, además de realizar una
aproximación real a través de un túnel de viento para validar los modelos anteriores.
Palabras clave: capacidad de dispersión, amenocoria, semillas, modelo de WALD, túnel de
viento.
Abstrac
Many literature have addressed the dispersive processes to understand them and
relate them to other ecological processes such as the relations between populations and
communities, evolutionary processes, colonizations, etc.
However, the parameterization and quantification of these processes is not very
abundant. This work will focus on dispersion through the wind vector, in other words,
anemomic dispersion and will attempt parameterization, measurement of the weight
variables in the process and prediction of the dispersive capacity of some annual herbaceous
species in the olive grove agro-ecosystem. This will be done by an approximation through
mechanistic models such as the WALD model or ballistic dispersion equations, in addition to
attempting to make a real approximation through a wind tunnel.
Keywords: dispersal capacity, anemochory, seeds, WALD model, wind tunnel.
1.Introducción.
1.1. Antecedentes.
1.1.1. Capacidad de dispersión, conceptos e importancia.
En la literatura científica encontramos una gran cantidad de estudios e
investigaciones con el fin de entender y explicar los procesos dispersivos de especies
vegetales, así como su importancia en relaciones ecológicas.
Luego para poder llegar a ese fin, es necesario definir dicho concepto de dispersión o
capacidad dispersiva para conseguir delimitar las componentes de peso que lo influencian.
La capacidad de dispersión se comprende por tanto como la capacidad de movilizar un
propágulo desde una fuente natal a través del espacio (Ronce, 2007).
Otros autores como como Tackenberg (2003) prefieren referirse a este concepto con
otra unidad de dispersión como la diáspora ya que no solo incluye la semilla con sus límites
morfológicos, sino también incluiría otras partes presentes en el momento de la dispersión
como frutos, vainas u otras partes vegetativas.
La dispersión es un proceso clave tal y como señala Jongejans (1999) para la
supervivencia, la distribución de poblaciones y comunidades vegetales, así como de sus
consecuencias en el flujo de genes. Es por esto, por lo que la parametrización y
cuantificación, como apuntan Harper (1977) y Nathan (2000), de la capacidad de dispersión
y las tasas de flujo de genes son de gran relevancia para analizar la biología, las dinámicas
o las estructuras poblacionales.
El conocimiento de la biología, concretamente de la fisiología, de la evolución y
diversidad existente apunta a que las plantas no son un elemento pasivo del proceso
dispersivo sino más bien piezas activas capaces de desarrollarse y adaptarse tanto a escala
temporal evolutiva, ecológica e incluso a periodos cortos de estrés para realizar lo más
efectiva posible la dispersión (Jongejans et al., 1999; Cain et al., 2000).
Son múltiples las adaptaciones logradas por las especies para dispersarse usando
vectores tanto bióticos, a través de especies zoológicas como abióticos como agua o viento
(Jongejans et al., 1999), este último será el vector de dispersión que se aborda en este
trabajo.
Pueden incluso dispersarse por múltiples agentes que intercambiándose pueden
aumentar las distancias totales como describe Nathan (2000). Esta combinación de agentes,
como señala el mismo autor, puede ocurrir en diferentes fases, señalando como fase 1 o
dispersión primaria a aquel movimiento desde la planta madre y como fase 2 o dispersión
secundaria al movimiento desde ubicaciones posteriores.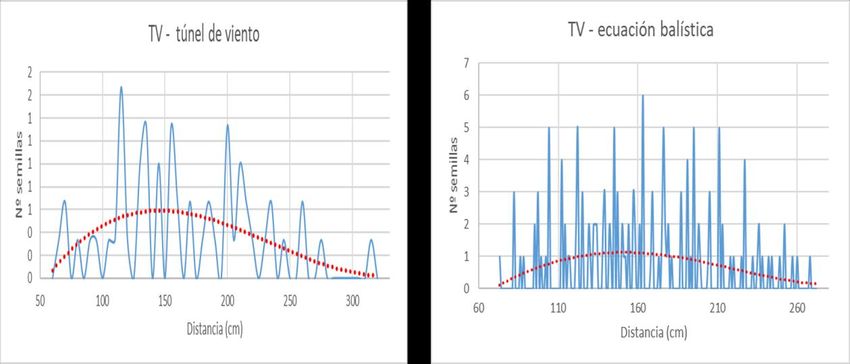
Como explica Nathan (2000) la complementación de estas dos fases reflejaría el
área potencial tanto de reclutamiento como de colonización, definida como “la capacidad de
establecer plántulas en sitios adecuados no ocupados previamente”, aportando información
sobre procesos de competencia, de depredación e intercambio genético.
Todas estas relaciones, aportados en la figura 1, son patrones determinados a partir
del patrón de dispersión de diásporas; a pesar de que estas relaciones y procesos han sido
descritos no se tienen suficientes datos cuantitativos (excepto las relaciones de denso-
dependencia con la hipótesis de Janzen-Connell), por lo para ofrecer una visión exacta,
actual y continuada es necesario cuantificar y parametrizarla (Harper ,1977; Okubo & Levin,
1989; Nathan et al., 2000).
Fig. 1. Las principales causas y consecuencias (en escalas de tiempo ecológicas) de los patrones
entre semillas en varias etapas (tomado de Nathan & Muller-Landau, 2000).
Este patrón general de dispersión aporta información sobre el potencial de
reclutamiento dependiente del patrón de disponibilidad de diásporas además del patrón de
éxito de establecimiento (Nathan & Muller-Landau, 2000). Este patrón de éxito de
establecimiento al igual que la capacidad de colonización tienen un concepto en común, el
grado de dispersión efectiva (Cain et al., 2000), todo ello relacionado con el contexto
espacial y ambiental, que será tratado más adelante.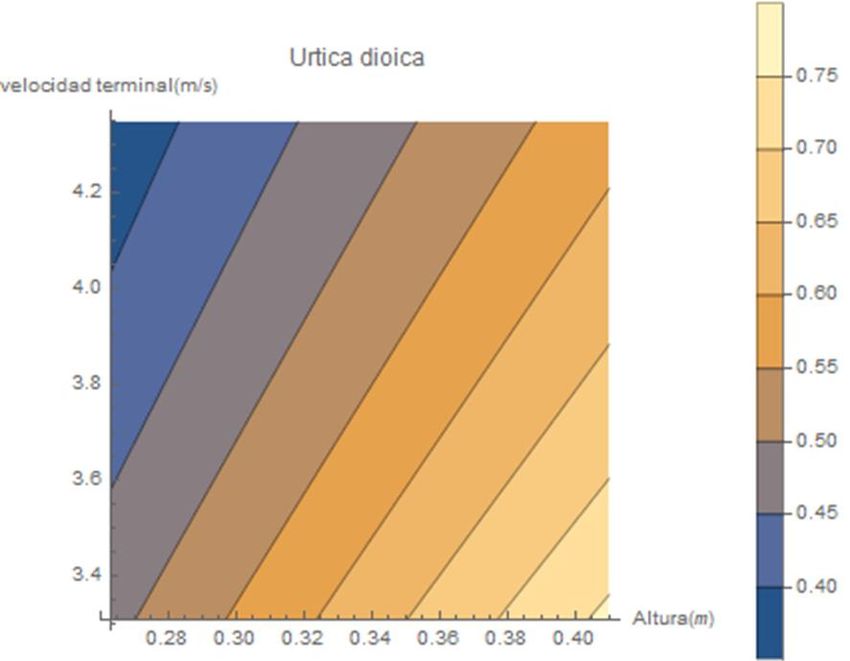
1.1.2. Parámetros influyentes en la dispersión anemócora.
Dentro de los diferentes vectores usados por las plantas ya mencionados
anteriormente, en este trabajo será objeto de estudio el viento, a la dispersión por este se le
denomina anemocoria.
Como también se ha explicado anteriormente, las plantas forman parte de la
dispersión de manera activa, por lo que las características de estas y de las semillas que
producen son de importancia para la dispersión. Esta variabilidad de morfologías para
realizar un correcto proceso, tiene origen en la filogenia; como las especies han optado por
estrategias o modificaciones de sus características para ello (Thompson et al., 2011).
En primer lugar, la altura de la planta y/o altura de abscisión de las diásporas.
Aquellas especies con mayor altura son capaces de dispersarse a mayor distancia que
especies de menor porte, siendo la variable de altura de la planta un componente con mayor
poder de explicación en la variación de distancia dispersada que la de la masa de la
diáspora en un análisis de componentes principales (Skarpaas et al., 2006; Thomson et al.,
2011; Johnson et al., 2019)
Por otro lado, la velocidad terminal es un parámetro resultante de la relación inversa
entre el peso de la diáspora y las características morfológicas de la misma en cuanto a
superficie expuesta al rozamiento o flujo de viento. Esta velocidad terminal se explica como
la velocidad que alcanza la diáspora al caer sin la influencia de ningún viento (Verkaar et al.,
1983; Augspurger y Franson 1987; Jongejans et al., 1999; Teller et al., 2014).
La velocidad terminal, como analiza Skarpaas (2006) a través una ecuación balística,
tiene una relación inversa con la altura de abscisión y la velocidad del viento en cuanto a
distancias de dispersión se refiere. Lo cual coincide con la propuesta de Greene (1993) en la
que, a menores velocidades terminales, mayor sería la dispersión final.
A su vez, este parámetro se propone como rasgo general de la especie ya que su
variación intraespecífica es menor que las variaciones de las componentes físicas del viento
(Jongejans et al., 1999; Horn et al., 2001).
En cuanto a los componentes individualmente, encontramos el peso el cual también
tiene de forma directa una relación negativa con la distancia de dispersión podría ser
resultado de la relación positiva de la masa de la semilla con la altura de abscisión
(Thompson & Rabinowitz, 1989; Moles et al., 2004).
La masa de la diáspora, por otra parte, puede tener consecuencias directas no solo
en la dispersión, sino también en aspectos fisiológicos en la planta como en la fotosíntesis,
ya que aquellos recursos destinados a aumentar esta masa no serán destinados a otros
procesos (Augspurger, 1988).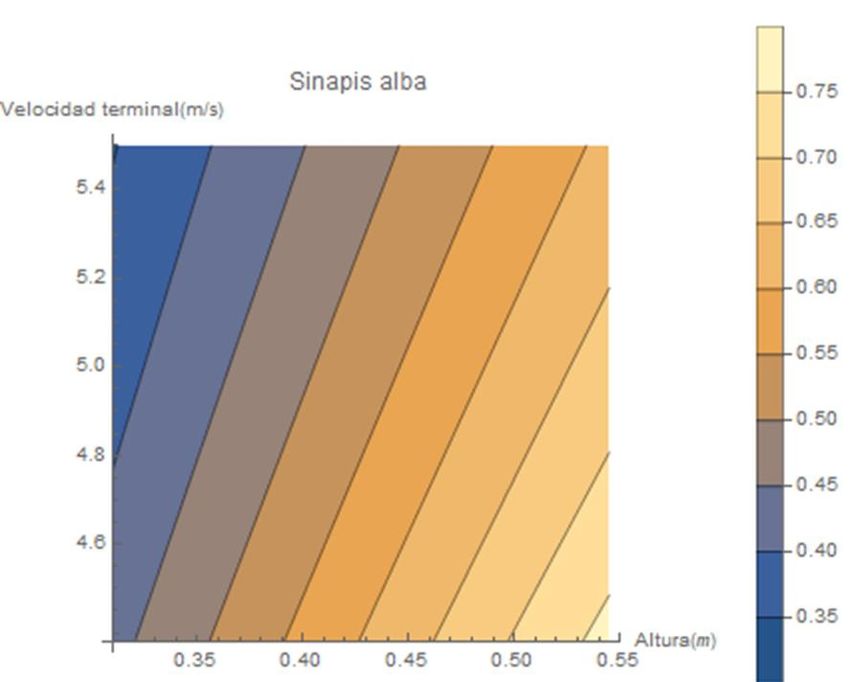
La morfología por su parte, tiene un efecto contrario al peso, por ello se han
desarrollado estructuras de vuelo para aumentar la superficie de exposición al viento para
aumentar la distancia de dispersión, encontrando semillas esponjosas, emplumadas o
aladas por ejemplo (Augspurger y Franson 1987; Jongejans & Telenius, 2001; Horn et al.,
2001; Seale et al., 2019).
Estas estructuras no solo ralentizan la caída por un mayor rozamiento, sino que
también como ha descrito Soons & Heil (2004), algunas especies plumadas de bajas
velocidades terminales son capaces de elevar su posición inicial con vientos verticales
ascendentes corrientes o por vientos ascendentes de convección como ocurre también en la
dispersión de algunos tipos de pólenes.
A pesar de esto, no son estas estructuras imprescindibles para una dispersión de
larga distancia ya que semillas generalistas o no especializadas en anemocoria también
pueden dispersarse por el viento (Tackenberg et al., 2003; Seale et a., 2020).
Por otro lado, además de las variables dependientes de la biología de la especie
botánica, la anemocoria lleva inevitablemente unida a ella influencias tanto del propio vector
de la dispersión como de otros factores abióticos.
Relacionado al viento encontramos tres características importantes. La primera de
ellas es la dirección de este, siendo los vientos horizontales y los verticales ascendentes los
más influyentes. Los vientos verticales ascendentes tienen un papel importante en la
abscisión; también presenta influencia unida a los vientos horizontales, que son la principal
componente del viento en términos de transporte de la diáspora, para retardar o incluso
elevar la semilla, aumentando así la distancia de dispersión (Jongejans 2001; Skarpaas et
al., 2006; Greene & Quesada, 2011).
La intensidad turbulenta del régimen de viento, por otro lado, es un parámetro
relacionado con la abscisión de la diáspora, ya que como expone Skarpaas (2006) a
velocidades bajas de viento, existe mayor número de semillas dispersadas en regímenes
turbulentos que laminares. Esta intensidad turbulenta se comprende como la variación de
velocidades en el régimen de viento (Katul et al., 2005; Teller et al., 2014).
Existe una correlación la cual indica que los vientos más potentes y extremos
conllevan una intensidad turbulenta mayor. Por lo cual, elevadas velocidades y turbulencias
del viento, mayor será el número de semillas liberadas y mayor la distancia a la cual se ha
dispersado.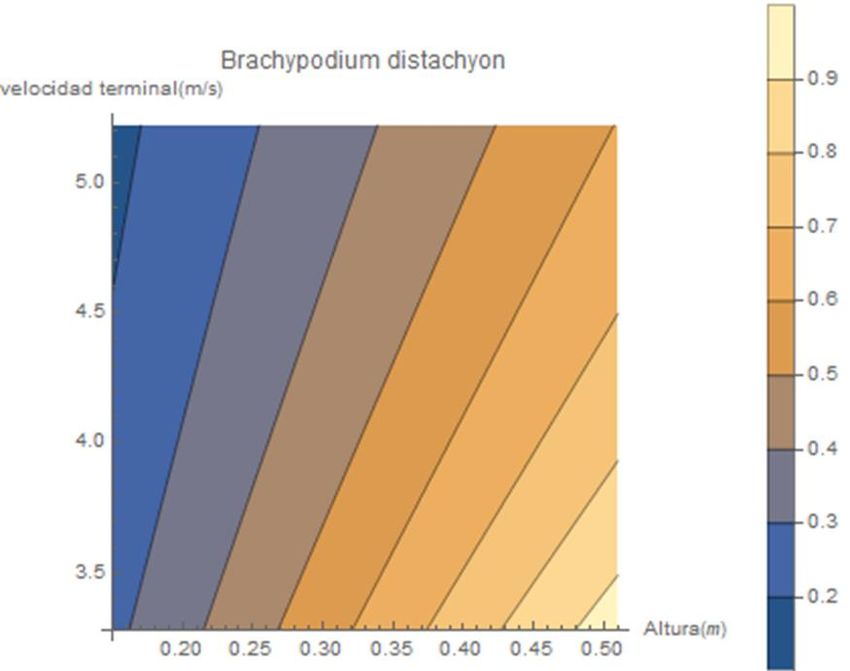
Además de las componentes del viento otros factores abióticos influyen en la
dispersión de diásporas, como por ejemplo la humedad relativa. El momento idóneo de la
abscisión no solo depende de los tiempos de formación de adecuada de la semilla, sino que,
también afectan condiciones ambientales como la humedad relativa (Greene & Johnson,
1989; Marchetto et al., 2010).
Además, otros condicionantes como la temperatura local, capaz de generar
corrientes de convección pueden influenciar el vuelo de semillas con velocidades terminales
bajas (Horn et al., 2001; Soons & Heil, 2004).
1.1.3. Modelos mecanicistas.
Los modelos mecanicistas son herramientas que relacionan variables
independientes, en el caso de la anemocoria, las características del viento, de la planta y de
la diáspora para predecir patrones de dispersión. Estas predicciones se realizan sobre un
sistema de estudio con datos reales de este, por lo que las variaciones y predicciones que
arroja serán muy cercanas a las existentes en el sistema (Nathan et al., 2001; Katul et al.,
2005).
En el caso que abordamos, estos modelos relacionan las variables con las distancias
de dispersión, que se construyen teóricamente al integrar la altura de lanzamiento de la
diáspora y la velocidad a la cual esta cae con la influencia de la velocidad horizontal del
viento en un intento de aproximarse a la velocidad de este (Katul et al., 2005). Estas
distancias se basan en ecuaciones balísticas que consiguen representar con acierto la moda
de las curvas de dispersión, aunque fallan en la representación de las colas, vinculadas con
eventos máximos o extremos de dispersión (Bullock & Clarke 2000).
En concreto, el modelo de WALD, es el modelo mecanicista de dispersión
anemócora sobre el que se basará este trabajo. Este nos aporta un rango de predicción y
comparación para evaluar como la variación de los valores de las variables, como la altura
de abscisión, la velocidad terminal, la velocidad horizontal del viento y la intensidad
turbulenta, tienen diferentes resultados finales en términos de capacidad dispersiva (Teller et
al., 2014).1.2. Justificación e hipótesis.
Como se ha descrito al inicio de este apartado, existe una gran cantidad de trabajos
e investigaciones con el fin de comprender los procesos de dispersión con el objetivo de
relacionar estos procesos con la ecología de poblaciones y comunidades, procesos de
colonización, procesos evolutivos de las especies, etc. Por el contrario, existe una literatura
limitada en cuanto a la parametrización, medición y predicción de la capacidad dispersiva.
Este es un trabajo que intenta cuantificar ciertos parámetros fisiológicos además de
la capacidad dispersiva de distintas especies anuales que forman parte de las cubiertas del
agroecosistema del olivar. Las especies herbáceas del olivar juegan un papel ecológico muy
importante en el control de la erosión entre otros servicios ecosistémicos. Así, en primer
lugar, analizaré las predicciones de distancias de dispersión usando tanto modelos
mecanicistas como el de WALD como ecuaciones balísticas, bajo la hipótesis nula de que
estos modelos son válidos para predecir distancias de dispersión. En segundo lugar, se
verificará la validez de los modelos mediante una aproximación a la realidad a través de un
túnel de viento.
2. Materiales y métodos.
2.1. Descripción de las especies estudiadas.
Especie Descripción Ecología
Arabidopsis thaliana -Hierba anual. Campos cultivados, baldíos
-Hojas en roseta basal. y bordes de caminos.
-Tallo erecto. Generalmente en
-Flores en racimo. ambientes secos.
-Fruto silicua con semillas
esfericas lisas.
Urtica dioica -Hierba anual. Arvense, ruderal y viaria.
-Hojas opuestas, pecioladas y Presente en suelos
dentadas. nitrificados.
-Inflorescencias terminales
bracteadas.
-Fruto aquenio.
Thymus vulgaris -Sufrútice. Lugares arbolados en
-Hojas opuestas elípticas y sustratos básicos. Clima
pecioladas. mediterráneo
-Inflorescencia capituliforme.
- Fruto tetranúcula.Conyza canadensis -Hierba anual. Ambientes ruderales y
-Tallo erecto. viarios.
-Hojas lanceoladas.
-Inflorescencia capituliforme.
-Fruto en aquenio con vilano.
Brachypodium -Hierba anual. Zonas húmedas tanto de
-Tallos con hojas alternas elevada altitud como zonas
distachyon
envainadoras. costeras.
-Inflorescencia en espiga.
-Fruto cariopsides.
Brachypodium stacei -Hierba anual. Pastizales secos de clima
-Tallos con hojas alternas mediterráneo.
envainadoras.
-Inflorescencia en espiga.
-Fruto cariopsides.
Brachypodium hybridum -Hierba anual. Pastizales secos de clima
-Tallos con hojas alternas mediterráneo, adaptable a
envainadoras. climas húmedos.
-Inflorescencia en espiga.
-Fruto cariopsides.
Sinapis alba -Hierba anual. Ambientes ruderales y
- Hojas grandes serradas y viarios.
pinnapartidas.
-Flores en racimo.
-Frutos en silicua con semillas
esféricas.
Ononis natrix -Hierba anual. Matorrales y pastizales
-Tallos muy ramificados. nitrófilos, ambientes
-Hojas trifoliadas. baldíos, ruderales y viarios.
- Unifloras agrupadas en el
ápice de los tallos.
- Fruto en legumbre
Tabla 1. Tabla descriptiva de las especies del estudio. (Información obtenida en
http://www.floraiberica.es/).
2.2. Localización y contexto.
La provincia de Jaén, está situada al sur de España, suroeste de Europa, esta
posición la expone tanto a vientos provenientes del Norte de África, Mediterráneo, Norte de
Europa o polares y vientos del Oeste. La península Ibérica se ve perturbada principalmente
por los vientos del oeste, caracterizados por ser húmedos y templados (Molina-Capel, 2015).
También se ve influenciada por los vientos de Levante (este) en los meses estivales
principalmente, estos son más cálidos y secos que el anterior. Los vientos polares rara vez
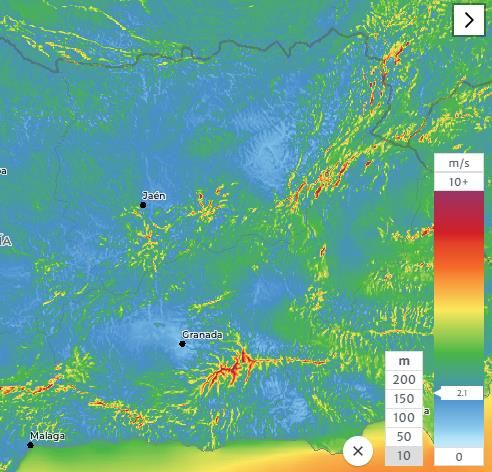
perturban la geografía peninsular (Muñoz, 1998).En cuanto a vientos más cercanos a la superficie (10m), que son los realmente
influyentes en la dispersión de las especies que estudiamos, observamos (figura 2) que la
provincia presenta generalmente tonos azulados y verdosos, correspondientes a
velocidades de entre 2,5-3,5 m/s, solo tomando esos colores pardos-rojizos en las cumbres
de cadenas montañosas.
Figura 2. Velocidades medias (m/s) a 10m. Fuente: globalwindatlas.info
Dentro de la provincia de Jaén las semillas fueron recolectadas en diferentes puntos
de zonas agrícolas de olivar del término municipal de Pegalajar, mientras que otras especies
fueron aportadas por el departamento de Biología Animal, Biología Vegetal y Ecología
(Tabla 2). Estas especies al estar localizadas en un agroecosistema tiene consecuencias
directas en la capacidad dispersiva y de colonización de las especies.
Especie Latitud Longitud Población
Arabidopsis thaliana - - Columbia (EEUU)
Urtica dioica 37.4359649 -3.390271 Pegalajar
Thymus vulgaris 37.7455772 -3.6085966 Pegalajar
Conyza canadensis 37.7455772 -3.6085966 Pegalajar
Brachypodium distachyon 38.3868998 -3.3802235 Hinojares
Brachypodium stacei 37.098183 -2.1239548 Sorbas
Brachypodium hybridum 37.757837 -3.797611 Jaén
Sinapis alba 37.7455772 -3.6085966 Pegalajar
Ononis natrix 37.7455772 -3.6085966 Pegalajar
Tabla 2. Tabla de coordenadas de localización de las especies estudiadas.La vegetación presente en un área cercana al individuo de estudio, puede suponer
tanto la competencia con los mismos en cuanto a la altura que deberían alcanzar para
conseguir una óptima dispersión (a mayor altura, mayor distancia de dispersión) como una
reducción en el impulso del flujo vertical y horizontal produciendo la deposición de las
diásporas (Figura 3) (Okin et al., 2006).
Figura 5. Relación entre densidad de plantas e interferencias en el flujo (Okin et al., 2006)
Las actividades agrícolas tradicionales junto a sus técnicas de laboreo eliminan las
posibles interferencias en el flujo debido a la eliminación de la cubierta vegetal, simplificando
el paisaje, mejorando así la actividad eólica y aumentando las distancias de dispersión (Okin
et al. 2006). La misma intensificación conlleva al deterioro del banco de semillas y
consecuentemente con la posibilidad de extinción de la biodiversidad que alberga (Soons &
Heil, 2002). A razón de esta reducción de biodiversidad e intensificación de la eliminación de
cubiertas produce la aparición de especies ruderales, viarias y de sucesión temprana como
es el caso de la mayor parte de las especies que estudiaremos, cuyo principal agente de
dispersión es el viento (Soons & Heil, 2004).
Las pequeñas poblaciones que quedan en parches no cultivados o baldíos, aportan
heterogeneidad al paisaje, convirtiéndose así en fuentes de colonización de las demás áreas
cultivadas no ocupadas debido al laboreo (Kaparinen et al., 2009; Teller et al., 2014).2.3. Fase de muestreo.
Para desarrollar la fase de muestreo se acudió a olivares cercanos del término
municipal de Pegalajar. Se detectaron aquellas fincas que presentaban parches de
vegetación de hierbas anuales para posteriormente tomando un margen de 15-20 metros
con respecto al camino para eliminar la influencia de especies puramente viarias y
ruderales. Dichos olivares presentaban en su mayaría un laboreo tradicional sin apenas
presencia de hierbas anuales propias del agroecosistema olivarero.
La recolección de semillas se realizó en el mes de septiembre y se basó en un
muestreo aleatorio de aquellas especies encontradas en los pequeños parches de
vegetación. Dichas semillas fueron almacenadas en sobres de papel transpirables para
evitar daños en la semilla además de anotar la especie correspondiente o en su defecto un
identificador general para su posterior identificación.
2.4. Germinación.
Para esta fase, se prepararon placas de petri con papel de filtro húmedo en su
interior sobre el cual se colocarían las semillas. Posteriormente las preparaciones se
expusieron durante una semana a condiciones de frio (en torno a 7ºC) y oscuridad en el
ambiente húmedo generado como simulación de las condiciones climáticas invernal,
necesarias para romper el periodo de latencia en algunas especies.
Tras este tiempo, se cambiaron las condiciones de la cámara a 22ºC con exposición
a la luz (300 µmolm-2s-1) con el fin de provocar la germinación de las semillas. En ambos
periodos, la cámara permaneció en condiciones normales de humedad relativa (alrededor
del 45%) y concentración de CO2 (400ppm).En cambio, Arabidopsis Thaliana, debido a las
dificultades que presenta su morfología, fue plantada directamente en maceta y esta, ya con
las semillas, era cubierta con un plástico transparente, proporcionando así las condiciones
óptimas para la germinación.
El tiempo de germinación de las semillas se anota a partir de la apreciación visual de
la radícula, mientras que el trasplante a una maceta se realiza tras el desarrollo de los
cotiledones.Figura 4. Preparaciones de placas de Petri con germinaciones de diferentes especies.
2.3. Mantenimiento y control de las plantas.
Las plantas germinadas fueron trasplantadas con ayuda de unas pinzas a macetas
cuyo sustrato fue previamente autoclavado para eliminar posibles germinaciones de semillas
o aparición de hongos presentes en el sustrato comercial.
Para el mantenimiento y control de las plantas fue necesaria una cámara de cultivo,
misma que para la fase germinativa, la cual permitiera controlar los parámetros ambientales
actuales que se precisaban para un correcto crecimiento de estas.
El mantenimiento de las plantas se basó en riegos periódicos cada 3-4 días. El riego
consistía en la adicción de la mayor parte del agua en las bandejas colocadas bajo las
macetas con la intención de obtener un crecimiento de las raíces hacia la base de la maceta
y obtener un cepellón mejor desarrollado. Además, con ayuda de un espurreador se
adicionaba agua en la parte aérea de la planta.Figura 6. Especies ya cultivadas en
Figura 5. Cámara de cultivo. macetas dentro de bandejas y con
cucharas usadas como identificadores.
En tres ocasiones, junto al agua de riego se añadió fertilizante Hoagland en las
bandejas para una absorción directa a través de las raíces, la disolución del fertilizante en
agua fue de 1/3 en probeta de un litro. La fertilización se realizó tras dos y tres meses
después de la germinación, en cada una de estas fertilizaciones se llevaron a cabo dos
aplicaciones con tres días de diferencia entre una y otra.
Debido a que algunas especies necesitan una señal de frío continuo para inducir la
floración (periodo de vernalización) y así completar su ciclo vital, se volvió a cambiar las
condiciones ambientales de la cámara durante cuatro semanas. La temperatura durante
este tiempo descendió a 7ºC para que cuando retornaran a las condiciones de 22ºC las
plantas iniciaran el proceso de floración.
Una vez desarrolladas las diferentes flores o inflorescencias y debido a la diversidad
de estrategias que polinización de cada una de las especies, se procedió al movimiento de
las macetas al jardín experimental para una polinización anemófila o zoófila.2.4. Análisis fisiológicos.
Después de los periodos de fertilización y de un correcto crecimiento, se analizaron
varios parámetros fisiológicos de las especies cultivadas con el fin de obtener información
de ellas e intentar relacionar los procesos dispersivos con la fisiología de la planta. Entre
ellos se realizó la medición de las curvas de saturación lumínica de la capacidad
fotosintética y se calculó el contenido relativo de agua (RWC) y del área foliar específica
(SLA).
2.4.1. Curvas de saturación lumínica de la capacidad fotosintética.
Para esta medición fue necesario un analizador fotosintético, en este caso el
instrumento LI-6800 (figura 7). El programa “Autolog_Light_response” fue el utilizado para
realizar dicha medición. El equipo fue programado con los mismos parámetros de humedad
relativa, concentración de CO2 y temperatura que la cámara de cultivo.
En cuanto a las diferentes intensidades lumínicas, se programaron los siguientes
valores en orden descendente (2000, 1800, 1600, 1400, 1200, 1000, 800, 600, 400, 200, 50,
0 ~ estos valores expresados en µmol m-² s-¹). La medida en cada valor estaba espaciada
en 2 minutos para la correcta adaptación de la planta a dicha intensidad.
Para cada especie, se repitió la medición a tres individuos diferentes con el fin de
obtener una media representativa de cada una de ellas.
Figura 7. Medición de las curvas de saturación lumínica de Brachypodium stacei con el LI_6800.Una vez terminadas las mediciones, los valores fueron manejados a través del
software RStudio a través de las librerías readxl, tidyverse ,ggpubr y ggplot2.
2.4.3. Área foliar específica.
Para la medición del área foliar específica y el porcentaje de agua de la hoja, se
necesitó el medidor LI-3000C (figura 8) además de una balanza analítica y una estufa de
secado. Para hallar dicho parámetro en primer lugar a la hoja seccionada del tallo se le mide
él área foliar con el LI-3000C, después con un pesa sustancias y una balanza analítica se
procede a medir la masa de la hoja, este valor representará el valor de peso fresco. A
continuación de dichas meciones se procede a meter las hojas en la estufa de secado
durante tres dias para una vez seca realizar otro pesado que supondrá el peso seco en las
siguientes fórmulas.
Ecuación 1. Ecuación de SLA Ecuación 2. Ecuación de RWC
Figura 8. Medidor de área LI-3000C.
2. 6. Caracterización de las diásporas.
Una vez terminados los procesos de floración, polinización y crecimiento de las
semillas se procedió a caracterizar las semillas de las diferentes especies tanto en peso
como en morfología. Para el peso se usó una balanza analítica introduciendo en
individualmente a las semillas en un pesa sustancias tarado con anterioridad.
Aquellas semillas con un peso igual o inferior al error intrumental de la balanza, como
es el caso de Arabidopsis thaliana, se pesaron en conjunto, posteriormente se procede al
conteo de semillas y la cantidad medida se divide por el número de semillas.Por otro lado, las dimensiones de las semillas fueron calculadas con una cámara de
alta precisión (figura 9) acoplada a un ordenador con el software DinoCapture 2.0. Se
tomaron tres tipos de medidas, longitud, ancho y profundidad. Cada una de estas medidas
fue repetida en 5 semillas por especie estudiada a excepción de Sinapis alba que fue
estudiada tanto en dimensiones como en capacidad dispersiva en dos tipos, con y sin vaina
debido a la apreciación de la dificultad de separar algunas semillas de la vaina
manualmente, por lo que el viento dificilmente podría realizar esta sepración.
Figura 10. Cámara de alta precisión Figura 11. Ejemplo de colocación de
acoplada a un ordenador la semilla para realizar las medidas
morfológicas.
2. 5. Modelo de WALD.
La finalidad de este trabajo, es el análisis de la capacidad dispersiva a través del
modelo mecanicista de WALD con las predicciones que aporta este, las distancias de
dispersión. Los modelos mecaniscistas como se ha explicado con anterioridad nos aportan
predicciones a partir de datos reales de las variables independientes del sistema. Es por ello
por lo que se realizaron distintos experimentos para la medición de cada una de las
variables del modelo de WALD.
Figura 12. Ecuación del modelo de WALD.
Siendo:A continuación, se describen los parámetros que aparecen en el modelo.
2.5.1. Parámetro L.
Este parámetro, representa la altura de abscisión de la semilla. Para hallar el rango
de las diferentes alturas a las cuales esa especie libera las semillas, así como de la media y
valores máximos y mínimos, se utilizó una regla para medir dichas alturas. Se emplearon
tres individuos de cada especie, así como de todas las alturas de abscisión de cado uno de
los individuos ya sea semillas individuales, espigas o vainas.
2.5.2. Parámetro F.
Para la velocidad terminal, correspondiente a este parámetro de gran importancia
para la dispersión como ya ha sido descrito, se utilizó un túnel vertical para la caída de las
diásporas sin influencia de ningún viento (Figura 12), además de una regla, un foco led y
una cámara de alta velocidad junto con el software Photron FASTCAM.
La posición de la cámara era opuesta a la del foco, entre ambos, el túnel vertical con
la regla con el fin de obtener una imagen nítida para en blanco y negro para diferenciar más
fácilmente la diáspora al caer. Gracias al conocimiento en imagen de la velocidad de
grabación de la cámara (fps), tiempo transcurrido de grabación y a las distancias aportadas
por la regla, se obtiene la velocidad en m/s.
Para cada una de las especies se tomaron 5 semillas diferentes con el fin de obtener
valores máximos, mínimos y medios, con excepción de Sinapis alba que como se comentó
con anterioridad, se dividió en dos tipos de semillas, con y sin vaina.
Figura 12. Túnel vertical usado para medir la velocidad terminal (F).2.5.3. Parámetro U.
Además de las variables dependientes de la planta y las propias diásporas, el
modelo incluye variables correspondientes al vector de dispersión. En este caso, U
corresponde a la velocidad horizontal del viento.
Los datos de este parámetro fueron medidos con un anemómetro en el túnel de
viento a diferentes intensidades de la potencia del ventilador concretamente al 50, 60, 70,
75, 80, 90 y 100% de esta potencia.
2.5.4. Parámetro φ.
La variable φ, representa la intensidad turbulenta, comprendida como la variación de
velocidades en el régimen de viento y, por tanto, es dependiente del parámetro anterior.
Para esta medición se usó un anemómetro de hilo caliente PCE con el software
Meeter realizando una medición en continuo de las distintas velocidades para un potencial
del ventilador concreto. Cada una de las potencias era medida tres veces, de los cuales se
realiza una medición media, posteriormente se normalizan dichas medias y se calculan las
desviaciones típicas de estos datos normalizados. Todo ello fue calculado con el software
MATLAB a partir de los datos del anemómetro PCE.
2.5.5. Parámetro r.
Relacionado con la distribución de las distancias de dispersión, es el parámetro que
nos predice el modelo de WALD. Para validar este modelo y comprobar que dichas
distancias son similares a la realidad, se realizó una doble aproximación, una en el túnel de
viento y otra a partir de una ecuación balística (r=L*U*F-1) descrita por Katul (2005) y que
incorpora el modelo de WALD como µ, determinado como un parámetro de posición (Teller
et al., 2014).
En la aproximación mediante la ecuación balística o µ, las distancias de dispersión
son un resultado de la multiplicación de la altura de abscisión y la velocidad horizontal del
viento, esto queda dividido por la velocidad terminal. Estas operaciones y curvas fueron
realizadas con el software Microsoft Excel.Para obtener los resultados de estas operaciones por tanto se usaron todas las
alturas de abscisión recogida en todas las especies, que darán variabilidad a los resultados
para una velocidad del viento determinada y una velocidad terminal resultante de las medias
obtenidas siendo una variable fija.
Se realizaron curvas kernels (gráficos de estimación de densidad) para las distintas
velocidades medidas y otra, efecto de la suma de las demás kernels, representativa de cada
una de las especies con excepción de Sinapis alba que obtenía dos, una con vaina y otra sin
ella.
Por otro lado, la aproximación a partir del túnel de viento se realizó introduciendo la
planta junto a la maceta dentro del túnel de viento, fuera de este se preparó sobre el suelo
una superficie blanca con marcaciones cada 5 centímetros (figura 13).
Para cada especie se sometía a una planta a 1 minutos de exposición con el túnel
3,3 m/s, posteriormente se procedía al conteo de semillas en las distintas mediciones
preparadas. Se retiraban dichas semillas medidas y se repetía la operación a 4,6 m/s y al
6,2 m/s. La media a las distintas velocidades de tres repeticiones a la misma velocidad con
plantas diferentes supondría el valor medio para la especie a esa velocidad, suma de las
tres repeticiones sería el resultado general para la especie.
Al igual que la ecuación balística o µ, los datos fueron analizados y tratados con
Microsoft Excel.
Figura 13. Túnel de viento y Figura 14. Arabidopsis thaliana como
superficie medida para el conteo de ejemplo de colocación de las plantas
diásporas. en el túnel de viento.Para Brachypodium distachyon, Brachypodium stacei, Brachypodium hybridum,
Sinapis alba y Ononis natrix, la intensidad turbulenta aportada por el túnel no era suficiente
para que las semillas o díasporas consiguieran liberarse de la planta por lo que las
mediciones de las distancias de dispersión de estas se realizaron exponiéndolas, ya
liberadas, directamente al flujo de viento.
Finalmente, para obtener las predicciones de las distancias de dispersión a partir del
modelo de WALD, se introdujeron los datos de las componentes bióticas y abióticas
calculadas con anterioridad en el software Wolfram Mathematica.
Definiendo en primer lugar el modelo, las variables y los distintos máximos, mínimos
y medias para cada una de las especies para obtener finalmente de cada especie curvas de
predicción de mínimas, máxima y medias en cuanto a distancias de dispersión, además de
un gráfico comparativo interespecífico.
Además, para ofrecer el rango completo de los valores obtenidos en cuanto a
velocidad terminal (F) y altura de abscisión (L).
2.6. Análisis multivariante.
A través del software RStudio se realizaron análisis multivariante utilizando distintos
datos tomados con anterioridad. Fueron necesarias las librerías lme4 (Bate D. et al., 20015),
Matrix (Bate D. & Maechler M., 2021), relaimpo (Grömping U., 2006), ggplot2 (Wickham H.,
2016) y readxl (Wickham H., 2019).3. Resultados y discusión.
3.1. Tasas de germinación.
A lo largo de la etapa de germinación se realizaron varios intentos de germinación de
las semillas recolectadas. Esto se debió a la baja germinabilidad de las especies recogidas
en campo, no así de las especies proporcionadas por el departamento de biología animal,
biología vegetal y ecología, tal y como aparece en la figura 15.
Estas tasas de germinabilidad tan bajas en las especies recolectadas pueden
deberse al tardío periodo de recolección, finales de septiembre, donde el ciclo de vida de
estas especies ya había concluido y las semillas presentes en la planta eran aquellas no
dispersadas y posiblemente en su mayoría no viables.
Figura 18. Tasas de germinación (en %) por especie.
3.2. Relación entre factores y distancias de dispersión.
Como se describió con anterioridad, existen múltiples relaciones de los diferentes
componentes bióticos y abióticos en cuanto a la capacidad de dispersión de las diferentes
especies.
Entre estas relaciones encontramos en primer lugar los parámetros propios de la
planta que tienen trascendencia en las distancias de dispersión, como son la altura de
abscisión y la velocidad terminal de la semilla (Tabla 3).Spearman’s ρ p-value
Altura 0,02453988 0,9464
Velocidad terminal -0,6829295 0,02951
Tabla 3. Resultados del análisis correlacional de Spearman de las distancias de dispersión
medias con la altura de abscisión y la velocidad terminal.
Mediante este análisis estadístico encontramos que la variación debida a los
parámetros bióticos, de los resultados obtenidos de las medias del parámetro r a través del
modelo de WALD, es explicada principalmente por la velocidad terminal de la especie,
teniendo entre estas dos variables una relación negativa muy significativa. Tal y como era
predicho por la literatura, la ecuación balística o parámetro µ del modelo de WALD, a
menores velocidades terminales mayores son las distancias de dispersión. Por otro lado, es
comprensible la insignificancia de la altura de abscisión con respecto a las variaciones de
las distancias a pesar de ser un parámetro importante en el modelo, esto es debido a la baja
variación de alturas medias de las especies estudiadas.
Obteniendo estos resultados, donde es muy significativa la velocidad terminal,
realizamos otro análisis comprobando que rasgo de la semilla era el más importante en
dicha velocidad (Tabla 4).
Spearman’s ρ p-value
Largo (mm) 0,1641345 0,6505
Ancho (mm) -0,321212 0,3677
Profundidad (mm) -0,369697 0,2956
Peso 0,7841982 0,007245
Tabla 4. Resultados del análisis correlacional de Spearman relacionando los rasgos de la diáspora
con la velocidad terminal.
Indagando en los resultados de este análisis, encontramos como rasgo más
significativo el peso de la diáspora, teniendo una correlación de Spearman de 0,78 con la
variabilidad de las velocidades terminales de las especies.
Otro factor importante en la velocidad terminal caracterizado por la literatura científica
es la superficie de la diáspora de exposición al flujo o, concretamente en este parámetro, la
superficie de la diáspora expuesta al rozamiento de la caída libre. Esta superficie, debido a
la aerodinámica de las semillas y observable en los videos necesarios para la obtención de
la velocidad terminal, es el producto del ancho y la profundidad de la diáspora.Ambos rasgos tienen una relación negativa (aunque no significativa en el estudio)
con la velocidad terminal, a mayores anchuras y profundidades de las semillas, obtenemos
una mayor superficie de exposición y, por ende, una menor velocidad terminal.
Por otro lado, tal y como Augspurger (1988) incluye en su trabajo, la masa de la
diáspora tiene consecuencias tanto en la dispersión como en funciones fisiológicas de la
planta, ya que hemos comprobado la influencia en la dispersión a través de la velocidad
terminal, importante parámetro de la dispersión anemócora, realizamos otro análisis para ver
si existen relaciones observables con los siguientes parámetros fisiológicos (Tabla 5).
Spearman’s ρ p-value
RWC 0,6785714 0,1095
SLA -0,8468812 0,0162
Fotosíntesis 0,8928571 0,0123
Tabla 5. Resultados análisis de correlación entre el peso de la diáspora y parámetros fisiológicos.
Como podemos observar en los resultados de este análisis, existen relaciones
significativas como describía Augspurger (1988). Concretamente observamos una relación
positiva con la capacidad fotosintética encontrando que las especies con una mayor media
en la masa de las semillas también presentan unos valores límites de fotosíntesis mayores.
Otra relación muy significativa la encontramos entre la masa de la semilla y el área
foliar específica, donde las especies con menor SLA (o mayor masa foliar), se corresponden
a elevadas masas. Por último, y con menor significancia de las anteriores, encontramos una
relación positiva entre el contenido de agua relativo en hoja (RWC) y la masa de la semilla.
Estas relaciones se podrían justificar mediante los requerimientos vitales de la
especie. Ya que esta especie necesita invertir estos recursos en aumentar la masa de la
diáspora, necesita tener una actividad fotosintética, una masa foliar y un contenido en agua
mayor para no comprometer otras actividades vitales de la planta.
Por tanto, se observan relaciones significativas de las distancias de dispersión con la
velocidad terminal y esta a su vez con la masa de la diáspora, además de la relación
significativa de la masa con la actividad fotosintética.Se realizó un nuevo análisis para ver si existía una relación directa de las distancias
de dispersión con la actividad fotosintética de las especies o por el contrario solo una
relación indirecta a través de los factores mencionados (figura 22).
Spearman’s ρ p-value
Fotosíntesis 0 1
RWC 0,1091089 0,8159
SLA 0 1
Tabla 6. Análisis multivariante relacionando las distancias medias de dispersión y
parámetros fisiológicos.
Como podemos observar, los diferentes valores fisiológicos de las especies no tienen
relación directa, aunque si indirectamente, con su capacidad de dispersión. Las relaciones
muy cercanas a cero de Spearman’s ρ, nos aportan información de que la variación de estos
parámetros no afecta a las distancias de dispersión de forma directa. Este mismo
diagnóstico se corrobora atendiendo a los valores de significancia, quedando
extremadamente lejanos a 0,05.3.3. Modelo de WALD, curvas k(r).
Para la elaboración de los rangos de las distancias de dispersión, de las diferentes especies,
se introdujeron tres tipos de datos. En primer lugar, las curvas de mínimos corresponden a
los mínimos cuantificados en la medición de cada parámetro dependiendo de su influencia
en la dispersión, es decir, la mínima altura de abscisión encontrada, la mayor velocidad
terminal, la menor velocidad horizontal del viento y la intensidad turbulenta menor.
Por otro lado, los máximos representan todo lo contrario, máximas alturas de abscisión y
velocidades terminales menor junto a los vientos más elevados experimentados y la
intensidad turbulenta mayor.
En cuanto a las curvas medias, suponen el promedio de todos los parámetros del modelo
para la especie.
Figura 16. Curvas k(r) de Sinapis alba con y sin vaina.Figura 17. Curvas k(r) de Arabidopsis thaliana, Urtica dioica, Thymus vulgaris y Conyza canadensis.
Figura 18. Curvas k(r) de Brachypodium distachyon, Brachypodium stacei, Brachypodium hybridum y Ononis natrix.
Como se ha comentado anteriormente, las curvas mínimas representan los valores
mínimos observados tanto en los individuos estudiados como de las características del flujo
de viento y las curvas máximas representan los valores máximos del modelo que
proporcionan una mayor distancia de dispersión.
En primer lugar, encontramos que estas curvas de máximos y mínimos se
encuentran dependiendo de la especie más alejadas o cercanas entre ellas. Esto se debe a
la propia variabilidad de los valores encontrados en los rasgos de la planta, ya que los
valores proporcionados al modelo correspondientes al viento son iguales para cada una de
las especies.
Encontramos, por tanto, que aquellas especies con mayor proximidad entre las
curvas de mínimos y máximos presentan una variabilidad en la altura de abscisión y
velocidad terminal menor. Aquellas otras especies, por el contrario, con un mayor
distanciamiento de estas curvas presentan una mayor variabilidad intraespecífica de los
mencionados rasgos.
Otro aspecto apreciable en las curvas K(r) lo encontramos en la amplitud de estas ya
que en todas las especies encontramos que la amplitud de las curvas máximas es mayor
que las de las mínimas. Esto nos aporta información sobre los procesos de dispersión
extremos como por ejemplo los de colonización. Un mayor espaciado o mayor distribución
real de las semillas proporciona a su vez una mayor posibilidad de encontrar un micrositio
óptico para la germinación de esta semilla además de encontrar menor competencia con las
semillas de la misma población.
Aquellas semillas dispersadas cercanas a la población fuente debido a la poca
variación espacial provocará conflictos de competencia además como está descrito en la
hipótesis de denso-dependencia de Janzen-Conell.
En cuanto a las curvas medias, también encontramos tendencias generales. En
estas, observamos un acercamiento más próximo a las curvas mínimas que a las máximas,
además de estar más próximas también en amplitud a las mínimas. Si realizáramos una
gráfica que uniera todas estas curvas obtendríamos una distribución sesgada positivamente,
es decir hacia la izquierda, como también encontramos en la curva de dispersión de la
hipótesis de Janzen-Conell.Podemos decir que la mayor parte de las semillas se agrupan en localidades más
cercanas a la población fuente, con una tendencia de alejarse lo máximo posible para
disminuir el efecto de madre nodriza además de ampliar su distribución espacial
disminuyendo las relaciones de competencia. Todo ello sin alejarse demasiado de la
población foco ya que cercana a ella encuentran condiciones ecológicas óptimas evitando la
necesidad de buscar un micrositio adecuado para la germinación y supervivencia. Por tanto,
probablemente encontremos la zona de reclutamiento cercana a las distancias descritas en
la curva k(r) de medias.
3.4. Modelo de Wald. Relaciones específicas entre la altura de abscisión, velocidad
terminal y distancia de dispersión.
Las siguientes figuras son el resultado de la relación entre la altura de abscisión y la
velocidad terminal utilizando el rango de valores de cada especie. Todo ello bajo las mismas
características del régimen de viento a través del modelo mecanicista de WALD.
Figura 19. Relación de la altura de abscisión y la velocidad terminal con la predicción de las
distancias de dispersión de Sinapis alba con y sin vaina.Figura 20. Relación de la altura de abscisión y la velocidad terminal con la predicción de las distancias de dispersión de
Arabidopsis thaliana, Urtica dioica, Thymus vulgaris y Conyza canadensis.Figura 21. Relación de la altura de abscisión y la velocidad terminal con la predicción de las distancias de dispersión de Brachypodium
distachyon, Brachypodium stacei, Bechaypodium hybridum, Ononis natrix.De acuerdo con los trabajos y estudios, mencionados en el primer apartado, acerca
de las variables propias de las especies botánicas influyentes en la capacidad de dispersión,
como son la altura y la velocidad terminal, los gráficos de las figuras de dispersión del
modelo aportan una información valiosa corroborando dicha literatura. Podemos observar
una tendencia a mayores distancias de dispersión a medida que encontramos tantas alturas
de abscisión más elevadas y velocidades terminales menores.
Estas relaciones son diferentes en cada especie como podemos apreciar en las
líneas que delimitan las distancias de dispersión con diferentes colores. A medida que estas
franjas toman una dirección hacia la horizontal, como ocurre en el gráfico de Conyza
canadensis, cada unidad de variación en la velocidad terminal supone grandes cambios en
las distancias de dispersión; no así con la variable de la altura donde en estos casos la
misma variación no trasciende en grandes variaciones de las distancias de dispersión.
En otros ejemplos, como Ononis natrix, las relaciones son totalmente diferentes. Las
variaciones en la velocidad terminal de las semillas provocan menores modificaciones en las
distancias de dispersión con respecto a las mismas variaciones de altura de abscisión.
Realizando otras comparaciones a través de estos gráficos, podemos observar la
importancia de las características propias de la planta. Como por ejemplo el peso, a través
de las especies de (Arabidopsis thaliana y Urtica dióica). Si escogemos una misma altura
para ambas especies observamos una gran diferencia en las distancias de dispersión, esta
es debida a la velocidad terminal y como ninguna de las dos especies presenta estructura
de vuelo específica para la anemocoria observamos que la diferencia reside en la masa de
la diáspora. La masa de Arabidopsis thaliana es significativamente menor que la de Urtica
dioica y por tanto Arabidopsis consigue mayores distancias de dispersión.
Otro parámetro importante, en la velocidad terminal, es la superficie expuesta al
rozamiento de la semilla o de exposición al flujo de viento; este también es apreciable a
través de estas gráficas, concretamente en Sinapis alba. Aquellas semillas que se dispersan
junto a la vaina que la envuelve presenta una velocidad terminal menor al presentar más
superficie de exposición al flujo en comparación a la masa que adquiere con esta. Al reducir
su velocidad terminal y ser dispersada a la misma altura que las que no disponen de dicha
vaina, las distancias de dispersión son mayores en las semillas que se dispersan junto a la
vaina.3.4. Comparación interespecífica de curvas k(r).
Figura 22. Comparación intraespecífica de las k(r) a partir de las medias de todos los parámetros.Un hecho observable, a través de la figura 29, es la similitud de la relación entre
distancia de dispersión y la amplitud del rango de dispersión como ocurría a nivel
intraespecífico. A nivel de comunidad observamos que aquellas especies con mayores
distancias de dispersión presenta a su vez mayores amplitudes en dicho rango para ocupar
una mayor área de colonización, aumentar la probabilidad de encontrar un micrositio
adecuado o disminuir relaciones de competencia.
En las especies estudiadas encontramos una menor variabilidad en el factor de altura
de abscisión que la variabilidad que encontramos en la velocidad terminal. Además, el
modelo para esta gráfica fue alimentado con igualdad de condiciones del régimen de viento
tanto en velocidad como en intensidad turbulenta, por lo que encontramos que la mayor
parte de la variabilidad de las distancias de dispersión es ocasionada, tal y como nos
informaba el análisis multivariante, por la velocidad terminal. Conyza canadensis, Thymus
vulgaris y Arabidopsis thaliana son precisamente las especies con valores más bajas en
dicho parámetro.
Otro hecho observado y descrito con anterioridad es el fenómeno de dispersión
utilizando dos tipos de diáspora en Sinapis alba, donde aquellas semillas que se dispersan
junto a la vaina consiguen realizar vuelos más largos que las que se dispersan únicamente
con la semilla, esto debido a un aumento en la superficie de exposición al flujo y por ende,
una menor velocidad terminal.
Sin embargo, tal y como se mencionaba en el primer apartado de este trabajo, las
estructuras específicas para la anemocoria no son imprescindibles para realizar largas
dispersiones. Claro es el ejemplo que encontramos en Arabidopsis thaliana, cuya semilla es
totalmente esférica, pero basa su baja velocidad terminal en su reducida masa.También puede leer

















































