Trabajo Fin de Autor/es Director/es Facultad de Veterinaria - Universidad de Zaragoza
←
→
Transcripción del contenido de la página
Si su navegador no muestra la página correctamente, lea el contenido de la página a continuación

Trabajo Fin de Grado en
Veterinaria
Mejora del crecimiento de corderos y la calidad de leche mediante el uso de
melatonina exógena
Improvement of lamb growth and milk quality through the use of exogenous
melatonin
Autor/es
Silvia Luis González
Director/es
José Alfonso Abecia Martínez
Facultad de Veterinaria
2020/2021
ÍNDICE
1. RESUMEN __________________________________________________________1
SUMMARY
2. INTRODUCCIÓN _____________________________________________________2
2.1. Melatonina ............................................................................................................................. 3
2.2. Características generales de la leche ovina .......................................................................... 10
2.3. Crecimiento de los corderos .................................................................................................. 13
2.4. Raza Rasa Aragonesa ........................................................................................................... 15
2.5. Justificación y objetivos ........................................................................................................ 17
3. MATERIAL Y MÉTODOS ______________________________________________17
3.1. Animales y procedimientos experimentales ........................................................................ 17
3.2. Análisis de leche ................................................................................................................... 19
3.3. Métodos de análisis estadístico ........................................................................................... 19
4. RESULTADOS ______________________________________________________20
4.1. Efecto de la melatonina sobre la composición y calidad de la leche .................................... 20
4.2. Efecto de la melatonina sobre el peso y el crecimiento de los corderos............................... 22
5. DISCUSIÓN ________________________________________________________24
5.1. Efecto de la melatonina sobre la composición y calidad de la leche .................................... 24
5.2. Efecto de la Melatonina sobre el peso y el crecimiento de los corderos .............................. 25
5.3. Reflexiones finales................................................................................................................ 27
6. CONCLUSIONES ____________________________________________________29
CONCLUSIONS
7. VALORACIÓN PERSONAL _____________________________________________30
8. BIBLIOGRAFÍA______________________________________________________31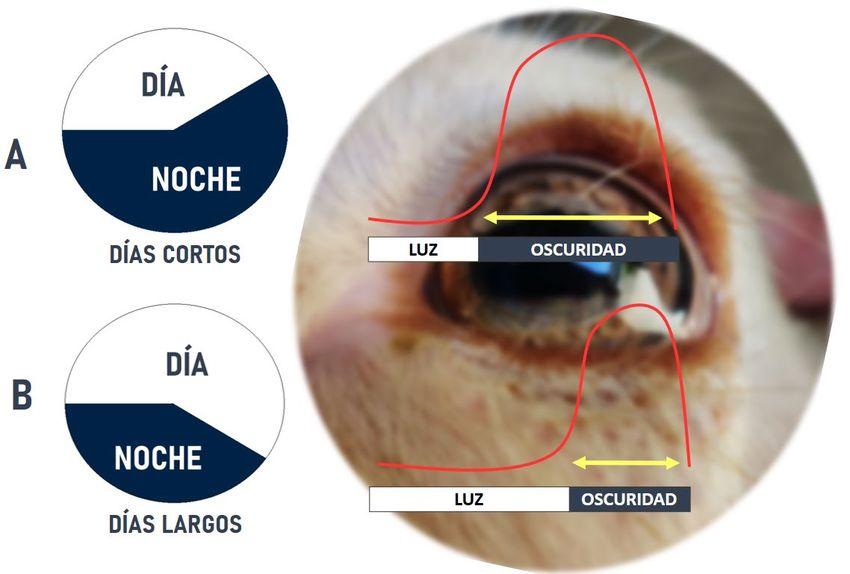
1. RESUMEN
Se diseñó un experimento para conocer y cuantificar los efectos que provoca la
melatonina exógena en ovejas y corderos implantados tras el parto, tanto sobre la calidad de la
leche de ovejas Rasa Aragonesa durante la lactancia como el crecimiento de sus corderos. Para
ello se contó con 55 corderos y sus madres (n=31) que recibieron (corderos: n=28; ovejas: n=15)
o no (corderos: n=27; ovejas: n=16) un implante subcutáneo de melatonina 24h después del
parto. Los corderos se pesaron (PV) semanalmente hasta el destete (6 semanas) y se calcularon
las ganancias medias diarias (GMD) en este periodo. Además, se tomaron muestras de leche a
los 15, 30 y 45 días después del parto. Con respecto al análisis de leche, el principal hallazgo fue
que en el día 45 de lactancia, la grasa de la leche y el contenido de sólidos totales fueron más
altos (P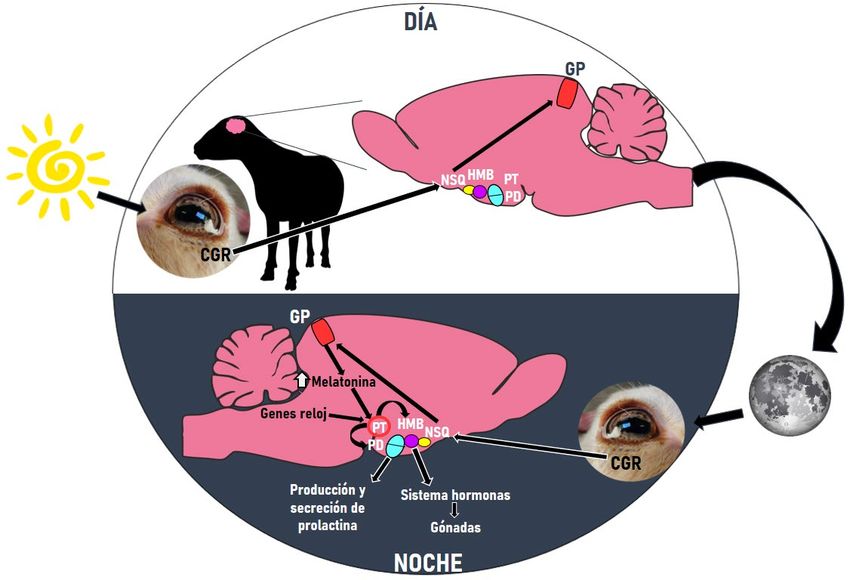
weaning (implanted: 13.61 ± 0.51; not implanted: 12.09 ± 0.57 kg) and ADG (implanted: 221.00
± 10.45; not implanted: 189.92 ± 12.44 g/d) than lambs raised by control ewes. In conclusion,
the melatonin treatment of the ewes at parturition increased the fat content of the milk and
induced a high growth rate in their lambs.
2. INTRODUCCIÓN
Algunas especies de rumiantes, como la ovina, muestran ciclos estacionales de
reproducción. Los animales de vida salvaje exhiben comportamiento reproductivo en otoño
(Abecia et al., 2012), que unido a un periodo de gestación de cinco meses, promueve que estos
nacimientos de corderos se den durante la primavera (Pool et al., 2020). El objetivo es que los
nacimientos se produzcan en época óptima de recursos para aumentar la probabilidad de
supervivencia de la progenie, hecho que no se eliminó con la domesticación (Abecia, et al.,
2012). Aunque la domesticación puede modificar la expresión fisiológica de la estacionalidad, la
mayoría de los pequeños rumiantes domesticados la han mantenido (Manca et al., 2014).
Los cambios fisiológicos y conductuales suceden como respuesta adaptativa frente a
cambios o fluctuaciones ambientales. El reloj biológico endógeno de cada animal es el que
permite interpretar y actuar en respuesta, y la luz es el principal factor ambiental que modula y
regula procesos fisiológicos, particularmente aquellos que siguen un ritmo circadiano como el
sueño y la reproducción, mediante variaciones en la concentración de melatonina (Correa y
Fernández, 2017). De este modo, la reproducción y sucesión de los ciclos estrales en el ovino
comienza en respuesta a la reducción de horas de luz o duración del día en verano o principios
de otoño y, termina a finales del invierno o principio de primavera, entrando en anestro (Abecia
et al., 2012). Esto es más evidente y se ven más influenciados por el fotoperiodo las especies de
ungulados que se encuentran en zonas del planeta de latitudes más altas, ya que cuanto más
cerca de la zona ecuatorial menor variación e influencia de la luz (Correa y Fernández, 2017).
Es necesario señalar que, si la reproducción es estacional, se genera un periodo de
partos que, teniendo en cuenta la producción, darán consecutivamente un periodo de venta de
animales de carne, y si los animales son lecheros, un patrón estacional de producción de leche
(Abecia et al., 2012). Por ello, para tener producción durante todo el año se ha tratado de
modificar los ciclos reproductivos.
La reproducción de los pequeños rumiantes puede controlarse a través de métodos que
incluyen, por un lado, los “métodos naturales”, como el control de la luz o la exposición a un
macho (el llamado “efecto macho”), y por otro, la administración de hormonas exógenas como
la progesterona o sus análogos (progestágenos), prostaglandinas, hormona liberadora de
gonadotropina (GnRH) y la melatonina. Sin embargo, mientras que la administración de
2
progesterona o sus análogos y las prostaglandinas, modificará la fase lútea del ciclo, la
melatonina actúa modificando la percepción del fotoperiodo y el patrón anual de reproducción
(Abecia et al., 2012).
2.1. Melatonina
La melatonina (N-acetil-5-metoxitriptamina) es una indolamina que fue descubierta por
primera vez en 1958 por Aron Lerner y sus colaboradores a partir del extracto de la glándula
pineal bovina (Lerner et al., 1958). Es una neurohormona fotoperiódica que se secreta en la
glándula pineal, se distribuye por la sangre e impulsa cambios estacionales de la actividad
reproductiva en la oveja. Que desempeñe un papel progonadal o antigonadal depende de si se
secreta en un patrón de día corto o largo, y si el animal es reproductor de días largos o cortos
(Bittman et al., 1983a).
Las concentraciones plasmáticas de melatonina son basales durante el día y altas
durante la noche (Figura 1), pero la señal que proporciona al eje hipotálamo-hipófisis-gonadal
sobre la época del año es la duración de la secreción nocturna. En este aspecto, la melatonina
es un estímulo progonadotrófico en los pequeños rumiantes ya que son reproductores de días
cortos (Abecia et al., 2012).
Figura 1. Las concentraciones plasmáticas de melatonina (línea roja) son basales durante el día y altas
durante la noche. En función de la duración del día, la liberación y duración de esta liberación cambia,
relacionándose los días cortos (A) (durante el invierno) con picos más largos, y los días largos (B) (durante
el verano) con picos cortos de melatonina. Al ser un reproductor de días cortos, el patrón de día corto con
picos más largos de melatonina es un estímulo gonadotrófico en los pequeños rumiantes (elaboración
propia).
3
En los mamíferos, la información de luz u oscuridad es percibida por las células
ganglionares de la retina cuyos axones viajan por un tracto nervioso y se proyectan en el núcleo
supraquiasmático (NSQ) del hipotálamo, considerado como el epicentro del reloj circadiano, ya
que a través de una compleja vía multisináptica se conecta con la glándula pineal (GP) y controla
el ritmo de síntesis y liberación de melatonina (Revel et al., 2007) (Figura 2). Por tanto, el patrón
rítmico se sincroniza a través de señales neuronales y hormonales mediante el NSQ (Li et al.,
2021).
La síntesis tiene lugar durante la oscuridad (Chowdhury et al., 2008). Las enzimas que
participan en ella son N-acetiltransferasa e hidroxiindol-O-metiltransferasa y, además de en la
GP, pueden encontrarse en el tracto gastrointestinal de varias especies de mamíferos (Bubenik,
2008). Durante la noche los niveles de melatonina en sangre son elevados y si cambia la duración
del día, cambia la liberación y duración de esta liberación, relacionándose los días largos
(durante el verano) con picos cortos de melatonina y, los días cortos (durante el invierno) con
picos más largos (Revel et al., 2007).
La actividad transcripcional en genes controlados por reloj circadiano se ve afectada por
las señales fotoperiódicas y de la melatonina que a su vez transmiten información a la pars
tuberalis hipofisaria (PT) y, es esta quién controla los niveles de moléculas funcionales que
transmiten mensajes y su salida neuroendocrina (Korf, 2018). La PT se encuentra
inmediatamente adyacente al hipotálamo medio basal (HMB) y es donde, en todos los
mamíferos, se encuentran los niveles más altos de receptores de melatonina (Dardente, 2012).
En las ovejas, se ha demostrado que solo se expresa un tipo de receptor de melatonina,
MT1 (Malpaux et al., 2001), por tanto, la melatonina actúa sobre los receptores de melatonina
MT1 de la PT, controlando de este modo su actividad y, la PT envía señales de salida a través de
vías retrógradas y anterógradas. La vía retrógrada conecta con el HMB y controla las funciones
reproductivas que, a través de la TSH y un sistema local de hormonas tiroideas en el hipotálamo
ejerce el control sobre la respuesta de las gónadas. Por otro lado, hay que mencionar que una
de las posibles vías anterógradas a destacar en ese estudio, es la conexión con la pars distalis
hipofisaria (PD) con el fin de controlar la producción y secreción de prolactina. En conclusión, la
PT transmite señales mediante un “cóctel” de moléculas mensajeras que actúan en otras áreas
cerebrales, desempeñando un papel esencial en la regulación de las funciones estacionales y
puede incluso ser la sede del reloj circanual (Korf, 2018). Además, los modelos actuales sugieren
que la melatonina es capaz de alterar la comunicación vascular entre las regiones funcionales
de la pituitaria, el infundíbulo, la PD y la PT, modificando la producción de hormonas
reproductivas (Hodson et al., 2012).
4Figura 2. La información de luz u oscuridad es percibida por las células ganglionares de la retina (CGR)
cuyos axones viajan por un tracto nervioso y se proyectan en el núcleo supraquiasmático (NSQ) del
hipotálamo. A continuación, a través de una compleja vía multisináptica se conecta con la glándula pineal
(GP) y controla el ritmo de síntesis y liberación de melatonina. Una vez llega la noche, la información de la
oscuridad, con la ayuda de las enzimas N-acetiltransferasa e hidroxiindol-O-metiltransferasa se sintetiza
la melatonina, y son elevados los niveles en sangre. Tanto la actividad transcripcional de los genes
controlados por el reloj circadiano como la señal de melatonina transmiten información a la pars tuberalis
hipofisaria (PT) donde se encuentran los niveles más altos de receptores de melatonina (MT1) y, es esta
quién controla los niveles de moléculas funcionales que transmiten mensajes y su salida neuroendocrina a
través de vías retrógradas y anterógradas. La vía retrógrada conecta con el hipotálamo mediobasal (HMB)
que a través de un sistema de hormonas controla las funciones fisiológicas. Una vía anterógrada es la
conexión con la pars distalis hipofisaria (PD) con el fin de controlar la producción y secreción de prolactina.
Con ambas vías se establece el eje neuroendocrino-gonadal (elaboración propia).
Es relevante destacar la función de la melatonina en la regulación de la secreción de
GnRH, ya que influye en la actividad reproductiva. Dentro de la compleja red de interneuronas,
se encuentran los terminales dopaminérgicos de la eminencia media que juegan un papel
fundamental en la modulación de la secreción de GnRH (Malpaux et al., 1997). Los pulsos de
GnRH se controlan por retroalimentación negativa tanto del estradiol como de la progesterona,
mientras que el aumento de GnRH se desencadena por la acción de retroalimentación positiva
5de altas concentraciones de estradiol al final de la fase folicular del ciclo ovárico (Nestor et al.,
2018).
Por último, con respecto al conjunto de acciones implicadas, se ha evidenciado que el
sistema Kiss1/GPR54 es responsable de controlar la reproducción a través del eje neural al
ajustar los niveles circulantes de gonadotropinas y esteroides sexuales, y que la melatonina
mediante acciones indirectas a este sistema transmite información sobre la duración del día al
eje neuroendocrino-gonadal (Revel et al., 2007). En función del patrón de secreción de
melatonina transmite información y gobierna la expresión del ARNm de KISS1 en el núcleo
arqueado (Smith, 2012).
Las oscilaciones constantes de la luz determinan la fluctuación de la concentración de
esta hormona proporcionando valiosa información al organismo. Estas fluctuaciones influyen
sobre el sistema endocrino y nervioso, manifestando cambios tanto a nivel molecular como
conductual (Correa y Fernández, 2017). Por lo tanto, los ritmos estacionales desempeñan un
papel importante en el control de las funciones corporales en casi todos los organismos vivos,
influyendo no solo sobre la reproducción, sino también sobre el metabolismo y las respuestas
inmunes. Se han atribuido importantes funciones a la PT, como la modulación de la respuesta al
estrés y las funciones inmunes (Korf, 2018). Además, han identificado receptores de melatonina
en diferentes lugares del cuerpo por lo que sus acciones ocurren a múltiples niveles, siendo el
eje hipotálamo-hipofisario la diana para modificar la percepción fotoperiódica del animal
(Abecia et al., 2012).
Los estudios más actuales contemplan que el efecto del temporizador de coincidencia
circadiana está asociado a cambios progresivos en todo el genoma, en el estado epigenético en
los promotores de genes estacionales clave, considerando que las interacciones entre el reloj
circadiano y las diferentes vías epigenéticas de la hipófisis son la base de este mecanismo
temporizador de los mamíferos (Wood et al., 2020).
Según Malpaux et al. (2001), se requiere un conjunto de mecanismos para que se
produzca respuesta a cambios en la duración del día. Deben detectarse tres tipos de cambios:
cantidad de melatonina que sobrepasa un umbral mínimo (noche frente a día); la duración de
esa cantidad de melatonina por encima del umbral (días largos frente a días cortos); y,
finalmente, la duración de la cantidad de melatonina en relación con la exposición previa a la
melatonina (duración del día en aumento frente a disminución). A pesar de esto, hay que tener
en cuenta que la diferencia entre la concentración diurna y nocturna es muy variable en cada
animal, aunque es altamente repetible en individuos con un fuerte determinismo genético
(Zarazaga et al., 1998). Además, cabe destacar que la respuesta al fotoperiodo no se basa
6únicamente en la duración absoluta del día, sino que también en la historia fotoperiódica que
hayan tenido previamente los animales (Robinson y Karsch, 1988).
Para controlar artificialmente el ciclo sexual en las ovejas, uno de los métodos que se
utiliza es la administración de melatonina exógena a través de implantes. Debido a que la
melatonina se libera por la noche, este procedimiento se utiliza para provocar una respuesta
similar a la de un día corto sin suprimir la secreción endógena. A pesar de que mantienen estas
elevadas cantidades de forma constante, no suprimen la secreción endógena de la hormona
durante la noche, esto explica que la melatonina exógena a través de implantes provoca una
respuesta de día corto por alargar la duración de la señal de melatonina (O'Callaghan et al.,
1991; Malpaux et al., 1997). Frente a los tratamientos hormonales convencionales para inducir
la ovulación en hembras, una de las ventajas de la utilización de melatonina exógena, es que
pueden usarse tanto en hembras como en machos, mejorando de este modo la fertilidad
resultante (Chemineau et al., 1992).
Los implantes que se comercializan contienen 18 mg de melatonina, y, mantienen
elevadas concentraciones plasmáticas durante más de 100 días (Forcada et al., 2002). Desde su
implantación hasta el comienzo de la actividad reproductiva pasan alrededor de 40 a 60 días
(AEMPS, 2009), se observa en la franja de estos días un cambio en la frecuencia del pulso de
GnRH y su consecutiva LH tras el inicio del tratamiento con melatonina (Malpaux et al., 1997).
Por otro lado, según el protocolo comercial del tratamiento los implantes se deben
colocar alrededor del día 45 después del parto por el estado fisiológico del animal, sin embargo,
Abecia et al. (2002) evidenciaron que los resultados reproductivos fueron mayores si se
colocaban alrededor del momento del parto. Del mismo modo, Forcada et al. (2002)
demostraron que la respuesta a los implantes de melatonina es mejor cuando se implantan
inmediatamente después del solsticio de invierno que alrededor del equinoccio de primavera
en ovejas con un período anestro estacional corto. Congruente con esto, se ha evidenciado
sistemáticamente que la transferencia de días cortos a días largos provoca la detención de la
ciclicidad en aproximadamente 30 días, es decir, muy rápidamente, mientras que la exposición
de ovejas en anestro o en días largos a días cortos provoca un retorno a la ciclicidad de forma
más lenta, entre 60 y 90 días (Bittman et al., 1983b).
Al comienzo, el uso de los implantes de melatonina se restringía al objetivo de adelantar
la temporada de reproducción, sin embargo, en la actualidad se ha incrementado el
conocimiento sobre otros usos prácticos en la mejora reproductiva de las ovejas, así como
aumentar su productividad tras la reproducción (Abecia et al., 2012).
En un metaanálisis, Palacín et al. (2011), encontraron que los implantes de melatonina
en genotipos mediterráneos aumentaron significativamente la fertilidad (+29%) y la fecundidad
7(+0,25 corderos adicionales por oveja implantada) de las ovejas. Esto es congruente con los
resultados de Abecia et al. (2007), en los que se concluyó que con los implantes se mejoran tanto
la fertilidad como el tamaño de la camada de las ovejas tratadas, aumentando el número de
corderos producidos entre un 15% y un 30% dependiendo de la raza, la granja y el momento de
la implantación. Además, se han obtenido resultados que indican que la melatonina influye en
el patrón de crecimiento de los folículos y en la capacidad esteroidogénica del cuerpo lúteo,
debido a que causa una disminución en la capacidad antioxidante del plasma y altera la dinámica
de crecimiento tanto del folículo como del cuerpo lúteo, y se interpreta que esto lleva a una
disminución en el número de ondas foliculares/ciclo y en el área media del cuerpo lúteo (Manca
et al., 2014).
El inicio de la pubertad define el rendimiento reproductivo de la oveja y, como tal, es un
factor crítico que influye en los resultados de la producción. Debido a que está sujeto a
limitaciones fotoperiódicas, además de metabólicas, se ha incorporado la melatonina para
inducir el inicio de la reproducción, y aunque debe ir unido a otros factores como el manejo
nutricional, factores sociales e incluso la selección genética, se ha observado que mejora la
fertilidad de las corderas tratadas con melatonina en comparación con las ovejas control y, da
como resultado características de producción similares a las de las ovejas maduras (Pool et al.,
2020).
En los últimos años se ha incrementado el interés por la predisposición genética a la
respuesta al tratamiento con melatonina. Mura et al. (2014) asociaron a la reproducción
estacional dos mutaciones silenciosas en uno de los principales genes: el receptor de melatonina
1A (MTNR1A). Identificaron que determinados genotipos exhiben tasas de fertilidad más
elevadas y una reducción en los días entre la introducción de carneros y el parto, concluyendo
que los polimorfismos en este gen en la raza de ovejas Sarda influyen en la reanudación
reproductiva de primavera. Por otro lado, Abecia et al. (2020b) confirmaron que determinados
polimorfismos en la secuencia del gen MTNR1A intervienen en el rendimiento reproductivo de
carneros Rasa Aragonesa jóvenes y adultos y, concluyeron que el genotipado podría ser un
procedimiento útil en el uso de los carneros en la ganadería ovina.
Por último, además de su importancia sobre la reproducción y los parámetros
productivos, esta molécula ha despertado interés por sus otras muchas funciones. Al tratarse de
una molécula lipofílica tiene libre acceso a todas las células, tejidos y órganos del cuerpo y, por
lo tanto, no requiere ningún receptor para llevar a cabo las acciones de eliminación de radicales
libres (Chowdhury et al., 2008). Una mayor secreción de melatonina se ha asociado con un mejor
estado oxidativo y una función cardiovascular alterada en varias especies de mamíferos (Lemley
y Vonnahme, 2017).
8En un estudio se evidenció que la administración de melatonina, en combinación con
hipotermia terapéutica, en bebés con encefalopatía hipóxico-isquémica mejoró su
supervivencia, redujo el estrés oxidativo y provocó un desarrollo neurológico favorable (Aly et
al., 2015). En otro estudio con bebés recién nacidos sépticos, se pretendía determinar los
cambios en el estado clínico y los niveles séricos de productos de peroxidación lipídica, mediante
la administración de melatonina, y, los resultados fueron consistentes: probaron una mejora
tanto del estado clínico como de los parámetros séricos (Gitto et al., 2001).
En consonancia con la investigación en humanos, en otro estudio en el que utilizaron
corderos como modelo animal, tanto la administración sistémica como transdérmica neonatal
redujeron los signos de neuropatología y encefalopatía asociados con la asfixia perinatal (Aridas
et al., 2018). Además, la administración posnatal de melatonina en corderos recién nacidos con
hipertensión pulmonar disminuye el remodelado vascular patológico y aumenta la angiogénesis,
promoviendo cambios funcionales y estructurales beneficiosos en la circulación pulmonar
(Astorga et al., 2018). También, se suplementó con cápsulas orales a ovejas merinas de gestación
simple y doble desde el día 80 de gestación hasta el parto (durante la última mitad de la
gestación), y se evidenció una mejora significativa en la supervivencia de los corderos gemelos
de segundo nacimiento debido a las acciones neuroprotectoras de la melatonina que aumentan
la tolerancia al parto prolongado (Flinn et al., 2020c).
En una revisión extensa sobre el uso de melatonina en ovejas y sus repercusiones en
corderos, se concluyó que: la suplementación durante la gestación mejora el flujo sanguíneo
uterino y posee potentes efectos antioxidantes que provoca una disminución del riesgo de
restricción del crecimiento intrauterino y anomalías neurológicas durante el desarrollo fetal,
protege el cerebro fetal de la hipoxia aguda pudiendo ser beneficiosa durante partos
prolongados y aumenta las reservas de tejido adiposo marrón que favorece la termorregulación
del cordero recién nacido (Flinn et al., 2020b).
Por último, además de sus repercusiones clínicas, asociadas en gran medida a su gran
poder antioxidante, también se ha relacionado con otros aspectos como su efecto sobre
folículos pilosos y conducta. Yang et al. (2019) demostraron que la melatonina promovió el inicio
y la maduración de los folículos secundarios posnatales y, aumentó su población y el efecto
beneficioso se mantuvo durante toda la vida. En definitiva, permitió un aumento de la
producción y mejoró la calidad de la fibra (diámetro reducido) asociado a una favorable
capacidad antioxidante y una supresión de la apoptosis. Por otro lado, Guesdon et al. (2013)
constataron que al retirar familiares conespecíficos se indujo respuestas de estrés conductual y
endocrino más fuertes durante la fase de luz que durante la fase de oscuridad. Estas diferencias
9en las respuestas de angustia entre el día y la noche demostraron el papel de la neurohormona
melatonina en la modulación de respuestas sociales estresantes en ovejas, mamíferos diurnos y
gregarios.
2.2. Características generales de la leche ovina
La leche de oveja se caracteriza por tener mayor cantidad de materia grasa y proteínas
que la leche de cabra y de vaca, y, generalmente también es mayor el contenido en lactosa.
Estas cantidades tan elevadas hace que cobre importancia en la producción de quesos y yogur
(FAO, 2021).
Para evaluar la potencialidad de los animales de ordeño, es muy importante tener en
cuenta la influencia de la etapa de lactancia ya que los datos deben ajustarse en función de esto.
En una revisión sobre el efecto del estado de lactancia sobre la composición de la leche en
diferentes razas de ovejas, se observó que los mayores contenidos de grasas y sólidos totales se
daban en la etapa tardía de la lactancia asociándose con una baja producción de leche. El
contenido de sólidos no grasos se mantuvo constante y se considera que la variación en los
sólidos totales se vio influenciada por la variación de la grasa. Por último, la secreción de lactosa
y agua son prácticamente constantes durante la lactancia (Assan, 2015).
Un factor relevante es la edad de las ovejas, las más jóvenes producen menos cantidad
de leche y menos rica en grasas, proteínas, caseína y proteínas séricas, y, según van
envejeciendo aumenta tanto la producción como el contenido de grasas y proteínas. La
alimentación es otro factor que se puede manipular para maximizar la producción de leche
durante las diferentes etapas de la lactancia (Assan, 2015). Suplementándose la dieta con grasa
protegida (by-pass) de aceite de palma en diferentes concentraciones a ovejas Lacaune
lactantes, se observó un efecto lineal positivo sobre el contenido de grasa de la leche. Los
animales que recibieron los niveles más altos de grasa protegida tuvieron una mayor producción
de leche durante la lactancia, y probablemente debido a esto demandaron mayores cantidades
de nutrientes para sintetizar leche más grasa, que provocó un menor aumento significativo de
peso corporal. A pesar de producir más, las proteínas, lactosa y los sólidos no grasos fueron más
bajos (Bianchi et al., 2018). Por último, analizándose registros entre 2007 y 2016 del control
mensual de ordeño de cabras Florida española, se observó que las cabras de parto simple
produjeron menos leche por lactancia y por día que las cabras de nacimiento múltiple. Las cabras
que tenían hermanos de camada machos produjeron más leche diaria y total, aunque con un
porcentaje de grasa más bajo que las cabras de parto único o que nacieron sin gemelos machos.
10Sin embargo, estas cabras que más producen, tuvieron una vida útil más corta que provocó que
tuvieran una menor producción de leche de por vida (Abecia et al., 2020c).
En la vaca lechera, se acepta que el sistema del reloj biológico está asociado con
alteraciones del metabolismo y hormonales; ambos son necesarios para iniciar y mantener la
lactancia (Casey y Plaut, 2012). Además, la regulación de este reloj circadiano controla perfiles
transcripcionales de genes circadianos centrales en la glándula mamaria (Hu et al., 2017) y,
recientemente se ha evidenciado que la expresión mamaria de un gen del reloj central (PER1) se
vio afectada por la exposición a los diferentes fotoperiodos y por el estado fisiológico de cabras
lecheras (Kalyesubula et al., 2021). Además, la fisiología circadiana domina la transición suave
de la gestación a la lactancia en las vacas lecheras (Li et al., 2021).
La estacionalidad reproductiva ovina da como resultado que el período de ordeño
natural comience durante los días largos, cuando disminuye la secreción de melatonina y
aumenta la de prolactina (Misztal et al., 1997), dependiendo los parámetros de producción de
leche tanto en ovejas como en cabras de las concentraciones de estas hormonas (Molik et al.,
2011).
La melatonina al provocar una respuesta similar a la de un día corto y aumentar sus
concentraciones plasmáticas basales, podría reducir los niveles de la prolactina que finalmente
provoca una disminución de la producción de leche (Abecia et al., 2005a). Las condiciones
ambientales y el fotoperiodo asociado a cambios en la secreción de prolactina en las ovejas
lactantes provocan la variación, principalmente en la cantidad de leche producida, así como en
la composición de la leche afectando a la síntesis de proteínas lácteas, grasas e
inmunoglobulinas (Molik et al., 2011). Se ha observado que la exposición a fotoperiodos cortos
durante el período seco estimula la lactancia posterior y se ha asociado al período prolongado
de secreción de melatonina por la glándula pineal (Avilés et al., 2019). En un estudio reciente
con cabras Saanen se corroboró el aumento de la producción de leche posterior en comparación
con el fotoperiodo de día largo. Además, la prolactina plasmática fue 20, 10 y 17 veces mayor a
las 3 semanas antes del parto, y 3 y 5 semanas después del parto respectivamente, en
fotoperiodos de días largos que en cabras sometidas a días cortos (Kalyesubula et al., 2021).
En las regiones de clima templado, las ovejas que paren en verano (días largos) producen
menos leche y tienen un periodo de lactancia más corto que las que paren naturalmente durante
el invierno (días cortos). Por ello, Misztal et al. (2018) diseñaron un estudio para observar cómo
influye la época del año en ovejas paridas en enero (días cortos), en junio (días largos) y, otro
grupo paridas en junio que recibieron implantes subcutáneos de melatonina un mes antes. Se
monitorizó tras el destete la producción de leche y la duración total de la lactancia. Se concluyó
que la producción de leche fue mayor en las paridas en enero y, el número de días de ordeño y
11la duración total de la lactancia fueron significativamente mayores en el grupo parido en enero
y el parido en junio implantado, que el grupo parido en junio sin tratamiento (Misztal et al.,
2018). Esto se explica debido al acortamiento de la señal de melatonina en primavera, y a que
el aumento de la concentración plasmática de prolactina en respuesta al estímulo de
amamantamiento y/o ordeño podría ser más potente en la estimulación de la síntesis y
liberación de leche (Lacasse et al., 2016). Por tanto, en respuesta a la prolongación de la señal
de melatonina durante el final del verano y el otoño, disminuyó la concentración de prolactina
y provocó un colapso en la productividad de la leche y acortó el período de lactancia (Misztal et
al., 2018).
Sin embargo, aunque la glándula mamaria de la oveja está igualmente preparada para
la síntesis y percepción de prolactina en condiciones de días cortos y largos dando como
consecuencia una producción similar en fotoperiodos crecientes y decrecientes, se ha
observado que esta similitud se produce solo durante el primer mes de lactación; a
continuación, las ovejas paridas en verano disminuyeron significativamente la producción de
leche. Además, durante la lactancia, las ovejas ordeñadas en el periodo de acortamiento del día
produjeron un 50% menos de leche que las ovejas ordeñadas durante días largos de primavera
y verano, sugiriendo que la melatonina en ovejas que se reproducen estacionalmente afecta a
la producción de leche posterior (Molik et al., 2007).
Son múltiples los factores que pueden influir en la producción de leche. En cabras se ha
establecido que intervienen tanto factores intrínsecos (la raza, la genética, el tamaño de la
camada y la edad) como extrínsecos (el sistema de producción, nutrición, la salud de la ubre, la
etapa de lactancia, la estación, la parición, el fotoperiodo, la duración de la lactancia, la
frecuencia de amamantamiento y ordeño) (Goetsch et al., 2011). Por lo tanto, para poder
comparar los resultados entre diferentes estudios, no solo se debe conocer cómo varían los
componentes a lo largo de toda la lactancia fisiológica, sino a qué factores y condiciones han
estado sometidos.
Por otro lado, estudios realizados indican que en las ovejas los parámetros de lactancia
y la composición química de la leche se ven influenciados por la duración del día y la señal de
melatonina, así como, en los parámetros bioquímicos de la leche influyen la temporada del parto
y el período en el que se inicia la lactancia (Molik et al., 2020).
En un experimento se dividieron 60 ovejas en 3 grupos: ovejas criadas en fotoperiodo
natural, criadas en fotoperiodo natural e implantadas con melatonina y ovejas expuestas
artificialmente a un fotoperiodo corto. Los resultados obtenidos mostraron que la
administración de melatonina exógena y la simulación de un fotoperiodo de día corto durante
el período estival tuvieron efectos significativos sobre los niveles de sólidos, proteínas, grasas y
12lactosa de la leche y sobre el contenido de ácidos grasos de la leche de oveja. Más
específicamente, la administración de implantes redujo el contenido medio de sólidos, la leche
del grupo control tenía un menor contenido de proteínas, la leche de ovejas expuestas a un
fotoperiodo corto y las implantadas contenían mayor nivel de grasa, siendo mayor la
concentración en las primeras. La melatonina disminuyó el nivel de lactosa después de 30 días
de lactancia y causó un deterioro significativo en el perfil de ácidos grasos de la leche (Molik et
al., 2011).
En la raza Churra se observó que el contenido y la composición de proteínas en la leche
no se vieron afectados por el número de corderos destetados y, congruente con otras
publicaciones, el contenido de grasa, proteína, caseína y proteína sérica, alcanzaron un mínimo
al mes de la lactancia, cuando la producción de leche era más alta, y aumentaron de forma
creciente a medida que avanzaba la lactancia (Othmane et al., 2002), hecho compatible con la
correlación negativa entre la producción de leche y la composición de la leche establecida en
extensa literatura (Bianchi et al., 2018). Además, se produjo un aumento en el recuento de
células somáticas que se asoció con una disminución de la producción de leche y de la lactosa.
En un estudio posterior, se estudiaron tres grupos de ovejas: paridas en febrero, en junio
y, en junio tratadas con implantes de melatonina. Los resultados mostraron que la mayor
cantidad de leche fue producida por ovejas paridas en febrero y la menor cantidad por animales
con partos en junio. Las ovejas paridas en febrero, además de una mayor producción de leche
en comparación con las ovejas de los otros grupos, se caracterizaron por una lactancia más
prolongada; sin embargo, la leche de las ovejas paridas en junio con o sin implante tuvo un
contenido significativamente mayor de materia seca, proteína y grasa. En todos los grupos
experimentales, el contenido de materia seca aumentó en el transcurso del período de lactancia
y, la composición química de la leche de las ovejas de lactación en junio, tanto con y sin
implante,en los primeros meses de ordeño fue similar a la de la leche de las ovejas de lactación
en febrero durante los últimos dos meses de ordeño (Molik et al., 2020).
2.3. Crecimiento de los corderos
Del mismo modo en que la reproducción de las ovejas se ve influenciada por el reloj
circadiano, también lo está el metabolismo y el crecimiento (Li et al., 2021), por ello, en la
actualidad se están presentando estudios en los que se observan mejoras en los rendimientos
de corderos nacidos de ovejas implantadas con melatonina durante la gestación,
evidenciándose un aumento de la supervivencia y el incremento significativo del peso al destete.
13Cabe destacar que los corderos durante las primeras semanas de vida tienen inmadurez
de la glándula pineal y por ello sus niveles endógenos de melatonina son arrítmicos y bajos
(Nowak et al., 1990), pero la suplementación materna puede administrar melatonina al feto
antes del nacimiento, debido a que es capaz de difundirse libremente a través de la placenta
ovina y la barrera hematoencefálica (Yellon y Longo, 1987; Aly et al., 2015). En un estudio de
1992, ya se concluía la importancia de mantener una transferencia rítmica diaria normal de
melatonina materna al feto durante la gestación (Kennaway et al., 1992). En la actualidad, se
piensa que el ritmo natural de la melatonina tiene una ventana de tiempo para ocurrir, ya que
los recién nacidos de varias especies carecen de la secreción rítmica de melatonina (Li et al.,
2021). En los corderos, una amplia exposición a melatonina en las primeras etapas de la vida
conduce a alteraciones obvias de la transición fetal a neonatal (Seron-Ferre et al., 2017).
Se ha contemplado que la administración de melatonina durante la gestación
desempeña un papel importante en la producción de tejido adiposo marrón (TAM) y en la
termorregulación del recién nacido (Seron-Ferre et al., 2015), ya que depósitos insuficientes de
TAM pueden contribuir a la muerte del cordero en las primeras 12 horas tras el nacimiento por
hipotermia, debido a la incapacidad de generar termogénesis sin escalofríos, lo que contribuye
a la inanición (Sales et al., 2017).
En ovejas con gestaciones simples y dobles, se implantó con melatonina exógena (uno
o dos implantes) a los 100 días de gestación, y se sacrificaron a los 140 días para extraer y pesar
la grasa perirrenal fetal; los resultados mostraron que aumentan especialmente en los fetos
dobles la deposición de TAM (Sales et al., 2017). Otros hallazgos de Seron-Ferre et al. (2015)
corroboran el papel de la melatonina materna durante la gestación en la cantidad y
funcionalidad de TAM del recién nacido. En este estudio, privaron crónicamente de melatonina
materna durante su último tercio de gestación y, se evidenció que estos corderos a los 5 días de
edad presentaban síntomas de hipotermia a diferencia de los recién nacidos de ovejas control
(Seron-Ferre et al., 2015). En consonancia, corderos recién nacidos de ovejas con secreción de
melatonina inhibida tenían menos tejido adiposo marrón y mostraban una respuesta de
temperatura cutánea exagerada a 4°C (Li et al., 2021).
Implantando melatonina a corderos de ovejas tanto pinealectomizadas como intactas
no se evidenció un efecto en el crecimiento por el tratamiento con melatonina (Kennaway y
Gilmore, 1984). Sin embargo, en una reciente publicación en la que se implantó a ovejas
diferente concentración de melatonina en los días de gestación 70-90, probaron que la
supervivencia del cordero se incrementó en ambos tratamientos a tres días posparto y se
mantuvo hasta el destete. Del mismo modo, las tasas de mortalidad relacionadas con el parto
14fueron mayores en los corderos de madres control. A pesar de esto, no se observaron efectos
en el peso al nacimiento del cordero ni a la tasa de crecimiento (Flinn et al., 2020a).
El aumento exponencial del crecimiento fetal durante la segunda mitad de gestación se
debe al crecimiento y vascularización del útero-placenta, y el intercambio de nutrientes y
metabolitos. En un estudio en ovino en el que se restringía el crecimiento intrauterino en
gestación media a tardía, se examinó el flujo sanguíneo uteroplacentario y el crecimiento fetal
si se administraba 5 mg/d de melatonina en la dieta y, se evidenció en las ovejas suplementadas
un aumento significativo del flujo sanguíneo arterial umbilical. Sin embargo, no se observaron
diferencias en el peso de los corderos al nacimiento (Lemley y Vonnahme, 2017).
Cabe destacar que, además de la posible participación de la melatonina, hay una amplia
gama de factores que interactúan con la tasa de crecimiento del cordero como: el peso vivo, la
condición corporal de la oveja, la capacidad de ordeño de la oveja, la calidad y cantidad del pasto
y/o alimentación y, la capacidad genética del cordero para crecer (Muir et al., 2003). Estos datos,
hacen pensar que es posible que haya una asociación entre los implantes de melatonina y el
crecimiento de los corderos, posiblemente asociado a múltiples factores; sin embargo, aún no
hay evidencias de que se produzca siquiera este efecto en ovino.
2.4. Raza Rasa Aragonesa
En el Ministerio de Agricultura, Pesca y Alimentación (MAPA) se caracteriza como una
raza autóctona de fomento que se encuentra de forma mayoritaria en la Comunidad Autónoma
de Aragón y agrupa en el año 2020, un total de 333.748 cabezas inscritas en el libro genealógico,
siendo la segunda española en censo tras la merina (MAPA, 2021).
Es una raza muy rústica que se adapta muy bien a diferentes medios geográficos siendo
capaz de aprovechar pastos muy duros (MAPA, 2021), permitiendo una valorización de recursos
renovables y sirve de base como una verdadera ganadería sostenible coordinada con el medio
rural (Sierra, 2000). El sistema de producción que llevan es semi-extensivo, se aprovechan los
recursos existentes con el pastoreo y se estabulan en periodos de parición hasta el destete
cambiándose su alimentación (cereales y forrajes) (MAPA, 2021).
Es de aptitud cárnica, caracterizándose por la producción de “Ternasco”; cordero con un
peso vivo (PV) medio de 22,5 kg que permite una adecuada finalización y una particular terneza
y sabor, cuya calidad ha permitido una indicación geográfica protegida (IGP) de “Ternasco de
Aragón” (MAPA, 2021).
La tasa de crecimiento en el primer mes de vida oscila entre 200 y 230 g/día en función
del sexo y forma de nacimiento, de los 30 a 90 días aumenta hasta 250-270 g, llegando a alcanzar
pesos finales de 24 a 27 kg (Sierra, 2000). Según la Asociación Nacional de Criadores de Ganado
15Ovino Selecto de Raza Rasa Aragonesa (ANGRA) los corderos se destetan con aproximadamente
45 días y se finalizan con piensos comerciales hasta los 75-90 días (ANGRA, 2021). Tras el
destete, los índices de trasformación se sitúan sobre 3,40 para los machos y 3,80 en las hembras,
ofreciendo rendimientos de canal entre 48,5-49,5% (Sierra, 2000).
Puede presentar la primera ovulación a los siete meses de edad, pero en las ganaderías
la primera cubrición se realiza mucho más tarde (Folch y Alabart, 2000). Esta precocidad sexual
permite cubriciones a los 7-9 meses de edad en corderas nacidas a primero de año y permitiendo
una amplia actividad sexual, que solo se ve disminuida en primavera (Sierra, 2000).
Al igual que el resto de ovinos, tienen mal o nulo comportamiento sexual en primavera,
sin embargo, esta raza presenta un buen comportamiento maternal todo el año (MAPA, 2021).
Tienen un anestro estacional corto, de poca intensidad, habiendo incluso un bajo porcentaje de
ovejas que se mantienen cíclicas y que pueden favorecer la respuesta al efecto macho (Folch y
Alabart, 2000). Según Forcada et al. (1992), tiene un período de anestroFigura 3. Oveja de la raza Rasa Aragonesa portadora del Gen GASE en la que la mayoría de partos
son gemelares. Estas ovejas se identifican con un crotal extra de color verde.
2.5. Justificación y objetivos
Teniendo en cuenta las evidencias, conocimientos y resultados de experimentos
expuestos anteriormente sobre la aplicación de melatonina exógena, sus efectos beneficiosos
en múltiples seres vivos, su relación con la reproducción ovina, así como su posible influencia en
parámetros productivos como el crecimiento de los corderos y la calidad en la leche, el objetivo
general de este trabajo consiste en conocer y cuantificar el efecto de los implantes de
melatonina tras el parto sobre el crecimiento de corderos de ovejas Rasa Aragonesa y la calidad
de su leche . A partir de este, se desarrollarán los siguientes objetivos específicos:
Determinar la composición química de la leche de la raza Rasa Aragonesa, relacionar los
parámetros que le confieren calidad e interpretar los resultados en función del grupo
experimental.
Determinar la evolución del crecimiento de corderos categorizados en diferentes grupos
experimentales e interpretar los resultados específicos.
Relacionar e integrar todos los resultados productivos obtenidos con el fin último de valorar
la funcionalidad de los implantes en el sector.
3. MATERIAL Y MÉTODOS
3.1. Animales y procedimientos experimentales
El experimento se llevó a cabo en el Servicio de Experimentación Animal de la
Universidad de Zaragoza (41° 40'N), bajo procedimientos aprobados por el Comité de Ética, y de
17acuerdo con el RD 53/2013 sobre protección de animales de experimentación, además de la
Directiva 2010/63 de la Unión Europea sobre la protección de animales utilizados con fines
experimentales y otros fines científicos.
Se utilizaron treinta y una ovejas Rasa Aragonesa de entre 4 y 6 años portadoras del gen
GASE. A lo largo del experimento las ovejas se encontraban en grupo, en condiciones de
iluminación natural, y en un corral con acceso a zona de parque al aire libre, por lo que las
condiciones climáticas eran las mismas en todas las hembras. Se alimentaron con paja ad libitum
y concentrado de acuerdo con el estado fisiológico, con una condición corporal adecuada para
la gestación. Se esquilaron a finales de abril – principios de mayo, y recibieron el manejo
sanitario habitual en ganadería ovina.
Se introdujeron machos de la misma raza durante un mes para monta natural a
principios de mayo de 2020. Se realizaron ecografías a los 60 días de gestación y se seleccionó
el lote de hembras gestantes que formarían parte del estudio. Las ovejas no fueron sometidas
a ninguna sincronización hormonal, por lo tanto, los partos se produjeron en una ventana de
aproximadamente 20 días (desde el 30 de septiembre hasta el 23 de octubre).
A medida que iban pariendo y con el objetivo de tener grupos homogéneos, se fueron
tratando (M; n=16) o no (C; n=15) con un implante subcutáneo de melatonina (18 mg, Melovine,
CEVA Salud Animal, Barcelona) (Figura 4) en la base de la oreja izquierda a las 24 horas del parto.
Figura 4. Implantes de melatonina (18 mg, Melovine, CEVA Salud Animal, Barcelona).
Los corderos nacidos de estas ovejas se dividieron a su vez en dos grupos homogéneos,
involucrando a 55 corderos (2 simples, 44 gemelos, 9 trillizos; 31 machos, 24 hembras) que
recibieron un implante de melatonina subcutáneo intraescapular (grupo m, n = 28; 16 machos,
12 hembras) o no (grupo c, n = 27; 15 machos, 12 hembras), también a las 24 horas posparto.
De este modo, los corderos se clasificaron en uno de cuatro grupos teniendo en cuenta los
grupos de tratamiento de su madre y el suyo propio (Cc, n = 15; Cm; n = 13; Mc, n = 12; Mm, n
= 15).
18Además, los corderos se pesaron con un dinamómetro digital al nacimiento y
semanalmente hasta el hasta el destete (6 semanas), calculándose así la GMD g/d). Por último,
también se tomaron muestras de leche a los 15, 30 y 45 días después del parto en todas las
ovejas. Tras la recogida de cada muestra de leche se procedió a su congelación para, una vez
finalizada la recogida, realizar el análisis de todas en una misma sesión.
3.2. Análisis de leche
Se analizó la composición química de las muestras de leche recogidas con un analizador
de leche (Lactoscan SAP+, Milkotronic LTD, Bulgaria), que se calibró para leche de oveja
siguiendo las instrucciones del fabricante. Se midió el contenido en grasa, proteína, lactosa,
sólidos no grasos, densidad, sólidos totales, pH, conductividad, contenido de sales y punto de
congelación.
Figura 5. Equipo de análisis de leche (Lactoscan SAP+).
3.3. Métodos de análisis estadístico
Con los datos obtenidos en el experimento se llevó a cabo un diseño factorial 2 x 2 x 2 en el
que el tratamiento con melatonina de hembras y corderos, y el sexo de los corderos fueron
efectos fijos.
Los efectos del tratamiento sobre el crecimiento del cordero, la GMD y la calidad de la leche
se evaluaron estadísticamente utilizando el GLM PROC (SPSS v.26) en un modelo que incluía
tratamiento con melatonina de los corderos, tratamiento con melatonina de sus madres,
sexo y su interacción.
También se utilizaron pruebas t de muestras pareadas para cada grupo materno con el fin
de detectar diferencias significativas en la calidad de la leche entre los días de muestreo de
leche.
19 Las diferencias se consideraron significativas a P
(P
1,5
a a b b c
% 1
0,5
0
-0,5
°C -1 a b
-1,5
S15 S30 S45 PC15 PC30 PC45
M C
Figura 6d. Porcentajes medios (± SEM) del contenido de sales (S) y punto de congelación (PC) de las
muestras de leche recolectadas en los días 15, 30 y 45 de lactancia de ovejas que recibieron (M) o no (C)
un implante de melatonina tras el parto (letras diferentes indican diferencias significativas P15 *
13 *
*
11
*
9
kg
7
5
3
Nac Sem1 Sem2 Sem3 Sem4 Sem5 Sem6
M C
Figura 7. Peso vivo semanal medio (± SEM) de corderos cuyas madres recibieron (M) o no (C) un implante
melatonina. (* indica diferencias significativas entre grupos en P5. DISCUSIÓN
5.1. Efecto de la melatonina sobre la composición y calidad de la leche
En este aspecto, el principal hallazgo del experimento fue el mayor contenido de grasa
de la leche al final de la lactancia en las ovejas tratadas, unido a que mientras el porcentaje se
redujo significativamente en las ovejas control, en las ovejas tratadas no se manifiesta esta
caída, manteniéndose los niveles más constantes desde el principio hasta el final de la lactancia.
Este efecto que se produce al implantarse melatonina exógena tras el parto en ovino, es hasta
donde sabemos, la primera evidencia descrita.
Algunos estudios previos se centraron en los efectos de los implantes de melatonina en
la segunda mitad de la gestación, o en la mitad del periodo de ordeño tanto en la producción de
leche y/o crecimiento de la descendencia. Los implantes de melatonina a mitad de la fase de
ordeño no tuvieron efecto significativo en la producción de leche en ovejas Lacaune y Assaf
(Abecia et al., 2005a). Del mismo modo, Hadi (2020) demostró que en ovejas Manchega y
Lacaune implantadas en el día 35 de lactancia no modificaba la producción y composición de la
leche. Sin embargo, un implante a cabras Cashmere en el día 50 de lactancia mostró un mayor
contenido de grasa que las cabras control; sin embargo, los rendimientos diarios, la proteína y
la lactosa eran mayores en las cabras control (Yang et al., 2019).
Que los niveles de proteína y lactosa en las ovejas control aumentaran de forma
significativa a medida que pasaba el tiempo de lactancia, puede indicar un comportamiento
normal y fisiológico de la glándula mamaria de reducir la cantidad de leche producida. Esto se
abala por la relación inversa entre producción de leche y contenido en proteína y lactosa descrita
en algunos estudios (Assan, 2015). Aunque la producción de leche no se midió en nuestro
experimento, estos resultados pueden indicar una menor producción de leche en el grupo
control debido a un efecto de dilución (Othmane et al., 2002), debido a la correlación entre
niveles bajos de proteína y lactosa y una alta producción de leche en las ovejas (Bianchi et al.,
2018).
Con respecto a la producción y calidad de leche, hay que tener en cuenta que las ovejas
se cubrieron de forma natural en época de anestro y comenzaron a parir en otoño, es decir, en
días cortos, tras el fotoperiodo largo. En este aspecto, Molik et al. (2011,2020) obtuvieron
resultados que demuestran que la lactancia tras un fotoperiodo de días cortos natural o artificial,
u ovejas sometidas a tratamiento con implantes de melatonina en periodo estival, provocan
tanto una mayor producción láctea como una mejor calidad de la composición de la leche.
Nuestros resultados obtenidos, viniendo de un comienzo de periodo decreciente en luz, unido
a la administración de melatonina al parto, hacen que cobren relevancia.
24También puede leer

















































